

 (Hemiculter leucisculus)、鲫(Carassius auratus)、短颌鲚(Coilia brachygnathus)转为似鱎(Pseudolaubuca sinensis)、
(Hemiculter leucisculus)、鲫(Carassius auratus)、短颌鲚(Coilia brachygnathus)转为似鱎(Pseudolaubuca sinensis)、淮河流域安徽段地处淮河中游,流经阜阳、六安、淮南、蚌埠、滁州等地,长约420 km,平水流量1 334 m3/s,平水面积6 200 hm2,流域面积669 000 hm2。该地区地貌特征为大平小不平,岗冲交错,水系复杂,除淮河干流外,两岸分布着一连串的天然与人工湖泊,是安徽省重要的湖泊分布区,鱼类资源十分丰富[1]。近年来,随着城市化进程的加快,对淮河流域生态系统的干扰日趋严重,工业废水、城市污水排入河道,使其自然调洪功能受到影响,部分河流功能出现退化现象,因此对淮河流域的水生生物状况尤其是湖泊湿地的水生生物状况进行调查研究十分重要[2]。
城西湖位于淮河中游南岸,是淮河流域重要的湖泊湿地,兼具调蓄洪水(蓄洪水位26.5 m,相应蓄洪面积517 km2)、维系生物多样性及支撑区域渔业发展的功能[3-4]。同时,该湖泊蓄洪为淮河流域储存了大量的淡水资源,在枯水期为其提供水资源,并为当地的生产生活提供了可靠的水资源保障[5]。城西湖具有“洪水一片、枯水一线”的水文特征,是研究禁渔初期鱼类群落响应与环境因子作用的理想区域[6]。2020年淮河流域实施禁捕政策,城西湖被纳入重点保护水域。目前关于城西湖的研究多在调洪蓄水规划方面,关于禁捕政策实施后城西湖鱼类群落结构与环境因子关系的相关调查和报道较少。在此背景下,本研究于2022—2024年的鱼类繁殖期(4—7月)和索饵期(8—11月)开展系统监测,以解析城西湖鱼类群落结构演变规律及其环境驱动机制,为淮河流域湖泊生态管理提供参考。
1 材料与方法
1.1 监测点设置
在城西湖(116°08′—116˚26′ E,32°10′—32°23′ N)进行分站点监测,每个站位分为3个小站点,每个站点进行2次捕捞收集样本,根据水域实际情况在城西湖共设置了2个站位,6个站点(图1)。
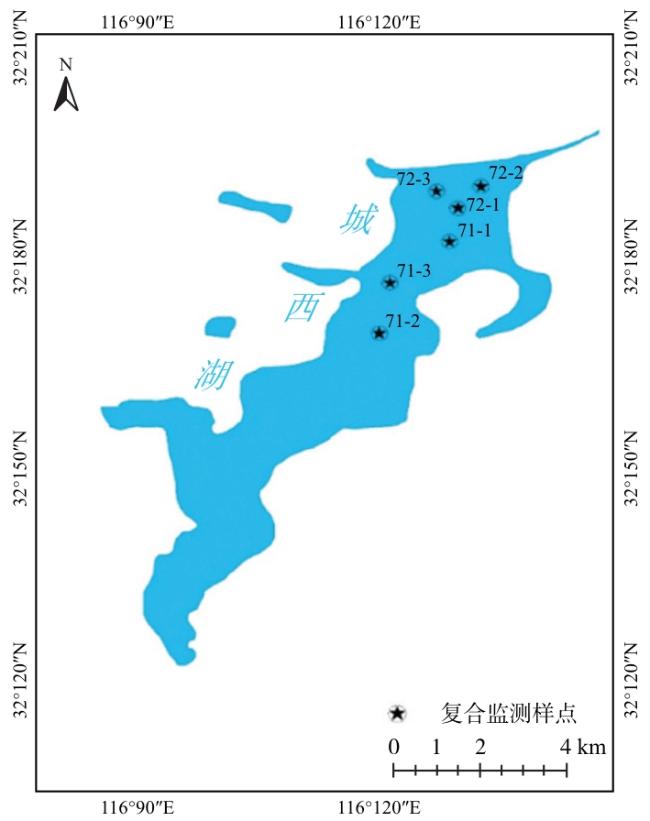
1.2 采样时间和采样方法
在2022—2024年鱼类繁殖期(4—7月)和索饵期(9—11月)分别监测1次,以确保监测的相对准确性。
渔获物测量工具:电子天平(FA2004N,上海精密科学仪器有限公司)测体重,游标卡尺(数显式,桂林量具刃具厂)量全长、体长等,标签、记录本记录信息。水质采样工具:有机玻璃采水器、水袋、便携式水质检测仪(YSI6000,赛莱默)。鱼类监测网具采用三层复合刺网(长×高为50 m×2 m,网眼直径2.0 cm)+(长×高为50 m×2 m,网眼直径6.0 cm)+(长×高为50 m×2 m,网眼直径10.0 cm)+(长×高为50 m×2 m,网眼直径14.0 cm)。和定制串联倒须笼壶(长×高×宽为18 m×0.33 m×0.45 m,网眼0.8 cm),于4:00—5:00放网,次日7:00—8:00收网。
表1 监测点水体理化指标 |
| 监测点水体理化指标 | 2022年F | 2022年S | 2023年F | 2023年S | 2024年F | 2024年S |
|---|---|---|---|---|---|---|
| 水温(WT)/℃ | 33.52±0.21 | 17.45±0.16 | 29.28±0.19 | 24.88±0.11 | 19.53±0.43 | 24.52±0.18 |
| 溶解氧(DO)/(mg/L) | 8.18±0.11 | 4.87±0.12 | 7.71±0.42 | 6.28±0.06 | 8.18±0.72 | 7.41±0.6 |
| 浊度(Tur)/NTU | 89.20±14.93 | 80.16±8.54 | 81.44±1.64 | 23.24±0.35 | 25.24±2.25 | 23.58±3.53 |
| 透明度(SD)/cm | 20.67±1.54 | 96.67±8.02 | 66.17±2.37 | 34.17±0.83 | 50.00±2.60 | 20.00±0 |
| 叶绿素a (Chla)/(mg/L) | 15.69±4.78 | 10.28±1.08 | 32.19±0.59 | 26.43±0.61 | 19.31±6.2 | 8.83±0.48 |
| 水深(H)/m | 0.92±0.95 | 2.75±0.11 | 1.83±0.08 | 1.80±0.07 | 1.83±0.08 | 1.83±0.08 |
| pH | 8.01±0.08 | 8.20±0.11 | 8.83±0.23 | 7.75±0.01 | 7.78±0.08 | 7.40±0.11 |
| 高锰酸盐指数(CODMn) | 9.67±0.59 | 5.33±0.31 | 7.57±0.37 | 6.56±0.15 | 6.54±0.25 | 7.41±0.16 |
| 总氮(TN)/(mg/L) | 0.92±0.07 | 1.73±0.11 | 1.47±0.11 | 1.42±0.04 | 1.15±0.06 | 0.91±0.18 |
| 溶解态总氮(DTN)/(mg/L) | 0.71±0.09 | 1.03±0.07 | 1.01±0.16 | 1.04±0.03 | 1.00±0.06 | 0.69±0.13 |
| 总磷(TP)/(mg/L) | 0.290±0.19 | 0.090±0.01 | 0.089±0.01 | 0.080±0 | 0.050±0.01 | 0.060±0.01 |
| 溶解态总磷(DTP)/(mg/L) | 0.050±0.02 | 0.040±0.01 | 0.045±0 | 0.350±0.20 | 0.022±0.01 | 0.010±0 |
| 磷酸盐(PO43-P)/(mg/L) | 0.013±0.003 | 0.018±0 | 0.047±0 | 0.050±0.01 | 0.002±0 | 0.010±0 |
| 铵态氮(NH4+-N)/(mg/L) | 0.315±0.04 | 0.498±0.02 | 0.763±0.05 | 0.480±0.03 | 0.160±0.05 | 0.120±0.02 |
| 亚硝酸盐氮(NO2--N)/(mg/L) | 0.010±0 | 0.035±0.01 | 0.001±0 | 0.012±0 | 0.002±0 | 0.010±0 |
|
1.3 数据处理与分析
1.3.1 群落优势种
群落优势种计算如式(1) 。
IRI=(N+W)×F
式中,N为某一种类的渔获物尾数占总渔获物的百分比;W为某一种类的生物量占总生物量的百分比;F为某一种类出现的站数占调查总站数的百分比。将IRI≥1 000的种类定义为优势种,100≤IRI<1 000的种类定义为重要种,10≤IRI<100的种类定义为常见种,IRI<10的种类定义为少见种。将IRI≥100的种类(包括重要种和优势种)视为群落主要物种[10]。
1.3.2 群落多样性
群落多样性计算如式(2 )~(5 )。
Shanoon多样性指数:H'=-
Pielou均匀度指数:E=
Simpson优势度指数:C=
Margale丰富度指数:DMG =
式中,Pi 是频度,Pi = ,S是类群数目,Ni 是第i类群的个体数,N是所有类群的总个体数[11]。
1.3.3 丰度/生物量曲线
利用丰度/生物量曲线(ABC曲线)可评估生态系统健康和群落结构,可用来分析研究区鱼类群落的受干扰程度。W值用于量化生态系统受干扰程度,计算如式(6) 。
W=
式中,S为物种数,Bi 和Ai 分别为生物量和数量的累计百分比。当W>0时,生物量曲线在丰度曲线上方,群落受干扰程度较小;当W<0时,生物量曲线在丰度曲线下方,群落受干扰程度大[12]。
1.3.4 相关性分析
在Origin 2024软件中对水温、溶解氧等水体理化指标进行皮尔逊相关性分析并绘图;使用Canoco 5.0软件对出现的优势种尾数生物量与环境因子进行约束排序探讨相关性,数据处理后采用冗余分析;使用蒙特卡洛检验结果显示第一轴和所有排序轴均有显著性差异(P<0.05),证明冗余分析结果有一定生物学意义。
1.3.5 群落结构相似性分析
2 结果与分析
2.1 鱼类种类组成
根据2022—2024年监测结果显示,研究区累计采集物种40种(2022年共采集物种29种,2023年采集物种28种,2024年共采集物种31种;表2),隶属于8目13科33属;甲壳类3种,其中本土物种2种,外来物种1种,为克氏原螯虾(Procambarus clarkii),鲤形目鱼类资源最为丰富(19属24种),占总数的60%;鲈形目2属4种,占总数的10%;鲇形目4属4种,占总数的10%。
表2 研究区鱼类群落组成 |
| 目 | 科 | 属 | 种 | 2022F | 2022S | 2023F | 2023S | 2024F | 2024S |
|---|---|---|---|---|---|---|---|---|---|
| 鲱形目 Clupeiformes | 鳀科 Engraulidae | 鲚属Coilia | 短颌鲚Coilia brachygnathus | + | + | + | + | + | + |
| 鲤形目 Cypriniformes | 鲤科 Cyprinidae | 鲢属Hypophthalmichthys | 鲢Hypophthalmichthys molitrix | + | + | + | + | + | |
| 似鱎属Toxabramis | 似鱎Toxabramis swinhonis | + | + | + | + | ||||
| 棒花鱼属Abbottina | 棒花鱼Abbottina rivularis | + | + | ||||||
| 鲌属Culter | 达氏鲌Culter dabryi | + | + | + | + | + | + | ||
| 蒙古鲌Culter mongolicus | + | + | + | + | + | + | |||
| 翘嘴鲌Culter alburnus | + | + | + | + | + | + | |||
| 原鲌属Cultrichthys | 红鳍原鲌Cultrichthys erythropterus | + | + | + | + | + | + | ||
| 䱗属Hemiculter | 䱗Hemiculter leucisculus | + | + | + | + | + | + | ||
| 贝氏䱗Hemiculter bleekeri | + | + | |||||||
| 鲂属Megalobrama | 鲂Megalobrama skolkovii | + | |||||||
| 团头鲂Megalobrama amblycephala | + | ||||||||
| 鲫属Carassius | 鲫Carassius auratus | + | + | + | + | + | + | ||
| 麦穗鱼属Pseudorasbora | 麦穗鱼Pseudorasbora parva | + | + | + | + | + | + | ||
| 鲤属Cyprinus | 鲤Cyprinus carpio | + | + | + | + | + | + | ||
| 鳊属Parabramis | 鳊Parabramis pekinensis | + | + | + | + | + | + | ||
| 似鳊属Pseudobrama | 似鳊Pseudobrama simony | + | + | + | + | + | + | ||
| 银鮈属Squalidus | 银鮈Squalidus argentatus | + | + | + | + | ||||
| 鱊属Acheilognathus | 兴凯鱊Acheilognathus chankaensis | + | + | + | |||||
| 大鳍鱊Acheilognathus macropterus | + | + | |||||||
| 鳙属Aristichthys | 鳙Aristichthys nobilis | + | + | + | + | + | + | ||
| 赤眼鳟属Squaliobarbus | 赤眼鳟Squaliobarbus curriculus | + | + | + | |||||
| 蛇鮈属Saurogobio | 蛇鮈Saurogobio dabryi | + | + | ||||||
| 鳑鲏属Rhodeus | 高体鳑鲏Rhodeus ocellatus | + | |||||||
| 似刺鳊鮈属 Paracanthobrama | 似刺鳊鮈 Paracanthobrama guichenoti | + | |||||||
| 鲈形目 Perciforms | 沙塘鳢科 Odontobutidae | 黄黝鱼属Micropercops | 小黄黝鱼Micropercops swinhonis | + | + | ||||
| 刺鳅科 Mastacembelidae | 中华刺鳅属Sinobdella | 中华刺鳅Sinobdella sinensis | + | + | + | ||||
| 泥鳅属Misgurnus | 泥鳅Misgurnus anguillicaudatus | + | + | ||||||
| 鮨科 Mastacembelidae | 鳜属Siniperca | 鳜Siniperca chuatsi | + | + | + | + | + | ||
| 虾虎鱼目 Gobiiformes | 虾虎鱼科 Gobiidae | 吻虾虎鱼属Rhinogobius | 子陵吻虾鯱Rhinogobius giurinus | + | + | + | + | + | + |
| 攀鲈目 Perciformes | 鳢科Channidae | 鳢属Channa | 乌鳢Channa argus | + | + | + | + | + | + |
| 丝足鲈科 Osphronemidae | 斗鱼属Macropodus | 圆尾斗鱼Macropodus ocellatus | + | ||||||
| 鲇形目 Siluriformes | 鲿科Bagridae | 黄颡鱼属Pelteobagrus | 光泽黄颡鱼Pelteobagrus nitidus | + | |||||
| 瓦氏黄颡鱼Pelteobagrus vachelli | + | ||||||||
| 黄颡鱼Pelteobagrus fulvidraco | + | + | + | + | |||||
| 鲇科Siluridae | 鲇属Silurus | 鲇Silurus asotus | + | ||||||
| 颌针鱼目 Beloniformes | 鱵科 Hemiramphidae | 下鱵属Hyporhamphus | 间下鱵鱼Hyporhamphus intermedius | + | |||||
| 十足目 Decapoda | 螯虾科Astacoidea | 原螯蟹属Procambarus | 克氏原螯虾Procambarus clarkii | + | |||||
| 长臂虾科 Palaemonidae | 白虾属Exopalaemon | 秀丽白虾Exopalaemon modestus | + | + | |||||
| 沼虾属Macrobrachium | 日本沼虾Macrobrachium nipponense | + | + | + | + | + | + |
|
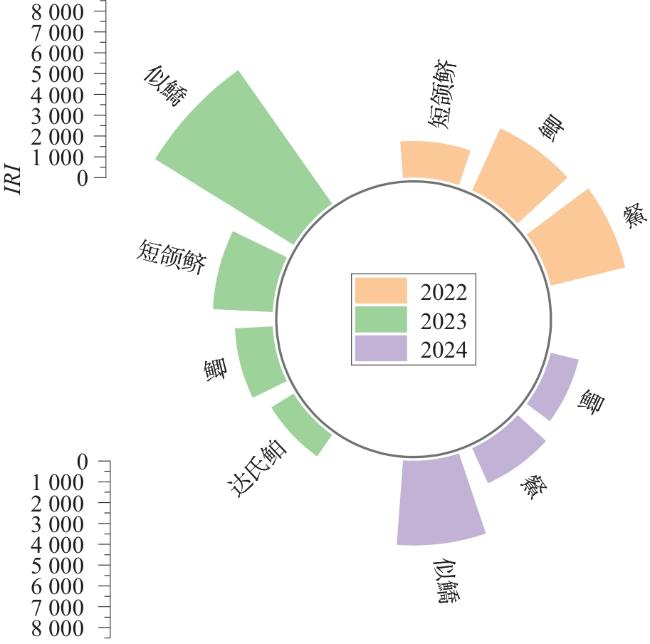
2.2 优势种变化
表3 研究区主要物种 |
| 时期 | 优势 种类 | 物种 | 数量占比/% | 重量占比/% | IRI |
|---|---|---|---|---|---|
| 2022年 | 优势种 | 䱗 | 28.68 | 18.057 | 3 699.98 |
| 鲫 | 6.42 | 26.239 | 3 266.13 | ||
| 短颌鲚 | 10.51 | 7.056 | 1 757.00 | ||
| 重要种 | 达氏鲌 | 5.19 | 4.350 | 954.28 | |
| 秀丽白虾 | 7.23 | 0.224 | 745.45 | ||
| 翘嘴鲌 | 6.05 | 3.369 | 588.85 | ||
| 似鳊 | 2.54 | 3.621 | 538.77 | ||
| 鲤 | 0.25 | 9.817 | 503.31 | ||
| 鳙 | 0.10 | 14.735 | 370.95 | ||
| 日本沼虾 | 30.73 | 1.696 | 135.13 | ||
| 2023年 | 优势种 | 似鱎 | 47.95 | 37.633 | 7 845.09 |
| 短颌鲚 | 20.71 | 20.062 | 2 887.96 | ||
| 鲫 | 3.42 | 15.672 | 1 829.75 | ||
| 达氏鲌 | 8.61 | 4.009 | 1 261.88 | ||
| 重要种 | 䱗 | 1.45 | 2.345 | 253.13 | |
| 似鳊 | 1.07 | 1.611 | 116.31 | ||
| 麦穗鱼 | 0.83 | 0.383 | 115.85 | ||
| 2024年 | 优势种 | 似鱎 | 57.05 | 24.20 | 4 062.39 |
| 䱗 | 21.32 | 15.78 | 1 854.96 | ||
| 鲫 | 4.60 | 23.19 | 1 389.89 | ||
| 重要种 | 似鳊 | 4.49 | 6.97 | 572.98 | |
| 子陵吻虾虎鱼 | 2.24 | 2.52 | 217.95 | ||
| 红鳍原鲌 | 2.39 | 1.55 | 180.63 | ||
| 达氏鲌 | 2.20 | 1.39 | 164.25 | ||
| 黄颡鱼 | 0.45 | 2.45 | 133.13 | ||
| 鲤 | 0.12 | 4.70 | 120.68 |
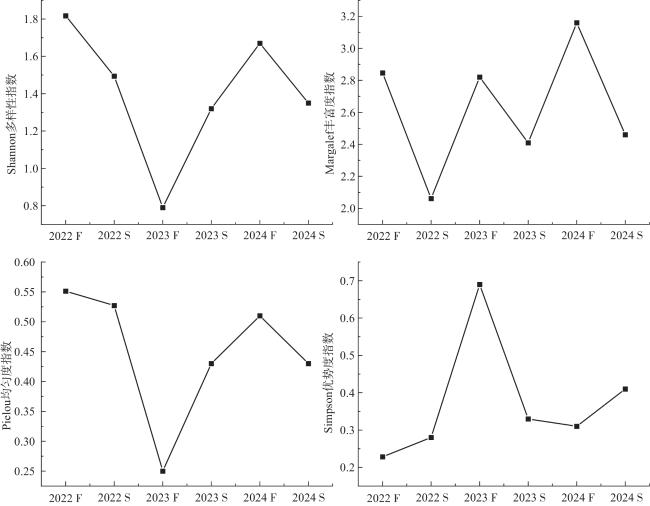
2.3 鱼类群落生物多样性指数变化分析
研究区鱼类群落的生物多样性在2022—2024年间呈现波动变化趋势。如图3所示,2022—2023年,生物多样性指数普遍下降,可能是由于环境因子变化或生物群落内部竞争加剧等,群落结构复杂性有所降低,优势种通过种内竞争和领域行为排斥其他物种,或是新迁入物种未能适应环境而数量减少,但整体仍处于相对较高水平,表明鱼类群落具备一定稳定性,此时期生态系统未出现明显改变。2023—2024年,湖泊生态系统有向好发展的趋势,鱼类栖息地环境得到改善,吸引更多鱼类种类定居,群落结构复杂性和稳定性增强。总体而言,研究区鱼类群落生物多样性在研究期间有一定改善,但仍需关注其动态变化,持续采取有效保护措施,以确保鱼类资源可持续利用和生态系统稳定。
2.4 丰度/生物量曲线分析
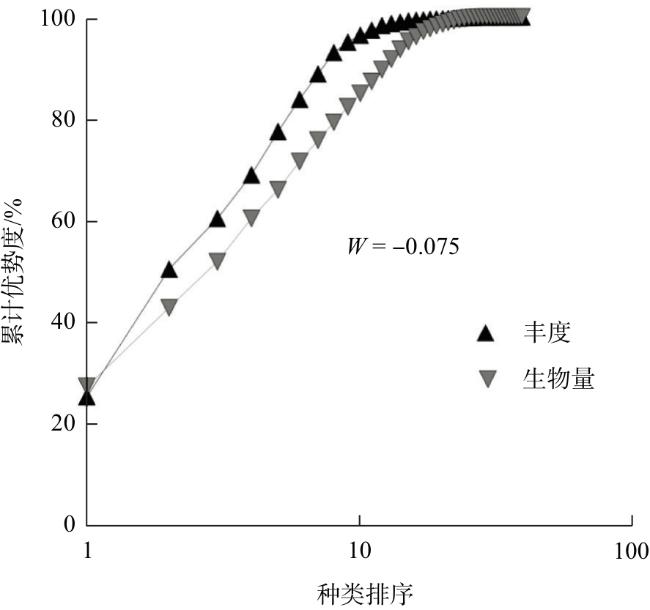
ABC曲线显示(图4),研究区鱼类丰度优势曲线总体上高于生物量优势曲线,W值为-0.075,表明禁捕初期城西湖鱼类群落受到的干扰较大,群落在向小型化、简单化发展。
2.5 环境因子与优势种变化的关系
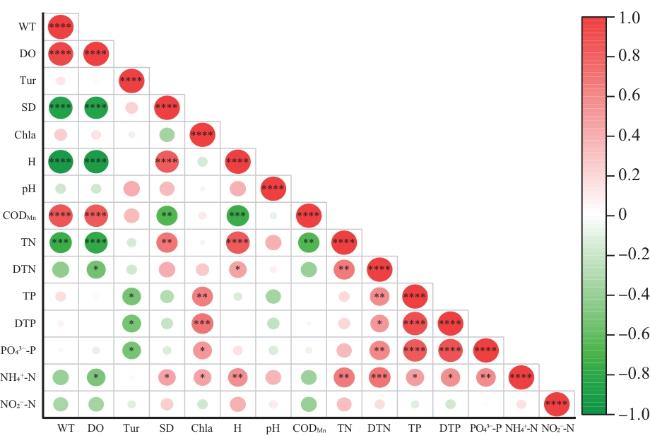
如图5所示,研究区水体理化指标中水温(WT)与高锰酸盐指数(CODMn)呈正相关(P≤0.000 1),反映了夏季微生物活性增强,会加速底泥有机质分解;总磷(TP)与叶绿素a(Chla)呈正相关(P≤0.01),表明磷是藻类增殖的关键限制因子,其藻类水华受磷负荷主导。
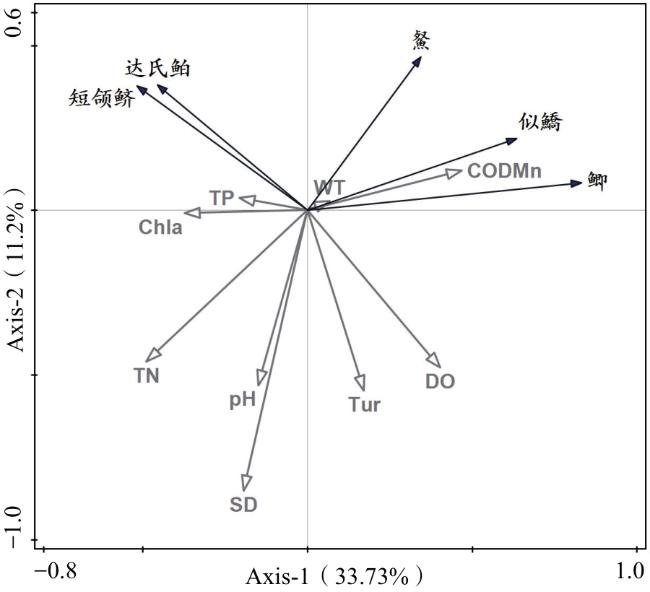
如表4所示,RDA统计图较好地说明了城西湖鱼类优势种变化与环境因子的关系。轴1(Axis 1)、轴2(Axis 2)上环境因子与物种的相关性分别是0.892 4和0.671 6,两轴共解释了优势种(44.93%)丰度数据的变化,其中物种与环境关系方差累计百分比为82.38%。
表4 城西湖鱼类丰度与环境因子蒙特卡洛分析 |
| 轴 | Axis 1 | Axis 2 | Axis 3 | Axis 4 |
|---|---|---|---|---|
| 特征值 | 0.337 3 | 0.112 0 | 0.077 7 | 0.016 9 |
| 物种累计方差百分比/% | 33.73 | 44.93 | 52.71 | 54.40 |
| 环境因子与物种相关性 | 0.892 4 | 0.671 6 | 0.666 1 | 0.404 2 |
| 物种与环境关系方差累计百分比/% | 61.84 | 82.38 | 96.63 | 99.73 |
冗余分析结果反映了5种优势种对湖泊各种环境因子的关系特点(图6),其中䱗、似鱎、鲫与水WT、CODMn呈正相关;TP、TN等其他因子呈负相关;达氏鲌、短颌鲚与TP、Chla呈正相关;与pH、溶解氧(DO)等其他因子呈负相关。
2.6 鱼类群落结构
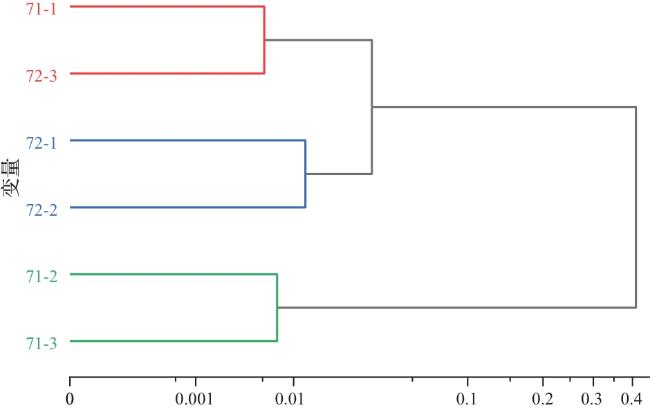
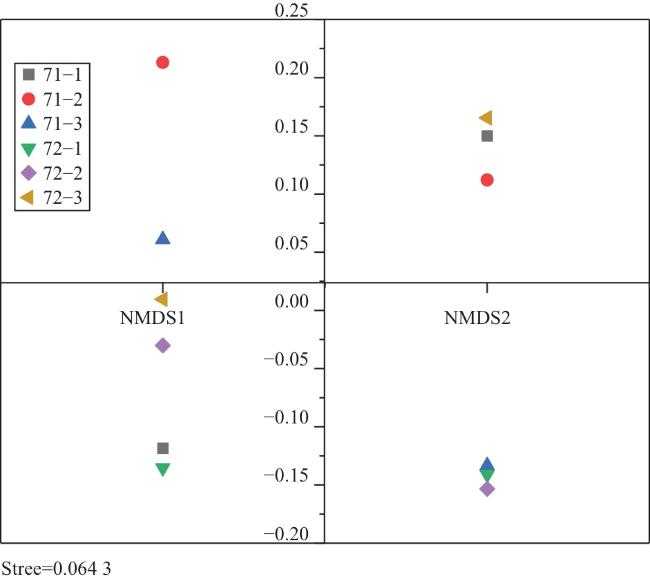
基于Bray-Curtis相似性(横坐标数值越小表示群落结构相似性越高)的聚类分析(图7)显示,站点间鱼类群落结构呈现明显的空间分组。具体而言,于图7横坐标刻度约0.4处,所有站点被划分为两个分离的类群:类群Ⅰ(71-2、71-3)与类群Ⅱ(71-1、72-1、72-2、72-3)。非度量多维尺度分析(NMDS)进一步验证了该分组模式的准确性,其胁迫系数Stress=0.064 3,小于0.1,结果可信(图8)。不同站点在二维排序空间中呈现与聚类分析一致的分组格局:71-2、71-3在空间中聚集,71-1、72-1、72-2、72-3 各自或成组分布,进一步验证了群落的空间分异。结合物种组成类群Ⅰ主要以䱗、似鱎等滤食性杂食性中上层鱼类为主;类群Ⅱ主要以鲫、达氏鲌、黄颡鱼等杂食性、肉食性中下层鱼类为主。上述结果揭示了城西湖鱼类群落结构在空间上的分异特征。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
城西湖作为淮河流域重要渔业水域,其生态服务功能不仅体现在优质水产品供给与渔民增收,更是水域生态文化建设和三产融合发展的关键载体,需统筹资源开发与保护管理[15]。
3.1 城西湖优势种变化及原因分析
3.1.1 短颌鲚的衰退
短颌鲚是一种洄游性鱼类,其繁殖过程依赖江湖之间的连通性,且对水质条件较为敏感[16]。本研究中,短颌鲚衰退的核心原因是城西湖2022年后水位调控措施阻断了其洄游路径,进而导致其失去了原有的产卵场所;另外,水质的富营养化可能抑制了幼鱼的存活(与DO负相关)。
3.1.2 达氏鲌的短暂优势
达氏鲌属于肉食性鱼类,主要以小型鱼类(䱗、似鱎)为食[17],且对DO要求较高,适宜生长环境为溶解氧含量大于5 mg/L。2023年,由于似鱎、鲫等小型鱼类数量的增加,为达氏鲌提供了丰富的食物资源,促使其在当年成为优势种;而到了2024年,尽管其索饵期溶解氧含量仍处于适宜范围(7.41 mg/L),但可能由于与其他物种竞争的影响,导致其退出了优势种行列。
3.1.3 似鱎的持续优势
似鱎是一种中上层滤食性鱼类,主要食物来源为浮游动物,对富营养化水体具有较强的适应能力[18]。2023年城西湖在鱼类繁殖期叶绿素a浓度较高,为32.19 mg/L,使浮游动物量增加,为似鱎种群的扩张提供了有力支持。其生态位的替代作用有效填补了短颌鲚衰退后的中上层空间,生物量占比从2022年的18%升至2024年的31%。
3.1.4 䱗的波动与鲫的稳定优势
3.2 环境因子对群落结构的驱动机制
冗余分析结果显示,第一轴(Axis1)解释了33.74%的物种与环境关系变异,其主要受TP和Chla驱动;第二轴(Axis2)解释了11.20%的变异,与DO和透明度(SD)相关。具体物种分布表现为鲫处于高TP、高Chla区域,显示出对富营养化环境的良好适应性;短颌鲚靠近高DO、高SD区域,依赖清洁水体生存,其在2024年的衰退与TP上升存在直接关联;似鱎与TP、Chla呈正相关,反映出作为滤食性鱼类,其在藻华期具有竞争优势。TP与Chla的协同作用是导致鱼类群落结构简化的核心因素,具体表现为耐污物种(鲫、似鱎)占据优势,而敏感种(短颌鲚)逐渐消退。
本研究表明,城西湖鱼类群落结构变化(如似鱎优势种更替)显著影响生态系统稳定,反映了鱼类资源动态既是湖泊演变的驱动因子,也是生态恢复成效的核心评价指标[22]。毛志刚等[21]在洪泽湖2017—2018 年鱼类群落调查中发现,其群落结构差异主要受水温、水位、水质污染和总氮调控,其中刀鲚(Coilia nasus)等洄游性鱼类的种群波动与TN浓度呈负相关,而鲫、似鱎等定居性鱼类则对水质污染的耐受性较强。这与本研究中,短颌鲚(同属鲚属)对清洁水体的依赖特性一致,体现出鲚属鱼类对水质污染的敏感性,但洪泽湖因兼具河流与湖泊双重水文特征,水位波动成为额外关键驱动因子,与本研究中城西湖以TP、Chla为主导的机制形成差异。
梁阳阳等[23]在巢湖(长江中下游重要湖泊)鱼类群落研究中指出,鱼类群落结构受捕捞强度、水深和洄游通道的影响,优势种为刀鲚和太湖新银鱼两种小型鱼类,占渔获物总量的96.84%,表明巢湖鱼类小型化明显。这一现象与城西湖鱼类群落结构高度相似,印证了禁捕背景下湖泊鱼类群落的共性演替规律,但巢湖因湖体面积大、水深差异明显,对鱼类群落分布的影响较大,而在城西湖中,水深对群落分布的影响未体现,反映出湖泊形态特征对驱动机制的调控作用。
3.3 城西湖鱼类群落结构
本研究发现,城西湖鱼类群落呈明显的营养级结构失衡,主要表现为小型杂食性鱼类(䱗、鲫)在群落中占据绝对优势(IRI>1 000),而大型肉食性及经济鱼类严重不足(IRI<100)。这一现象与罗思等[24]对淮河流域湖泊的群落退化特征研究结果相似。
3.3.1 过度捕捞的遗留效应
历史高强度捕捞选择性移除大型鱼类(如鳜、乌鳢),导致顶级捕食者缺失,小型鱼类失去天敌控制。类似现象在长江禁渔初期研究中被证实,王银平等[25]研究发现,禁渔前捕捞压力使长江下游鱼类个体小型化,群落生物量曲线显著低于丰度曲线(W<0)。
3.3.2 栖息地单一化限制
围垦和闸坝建设阻隔了江湖洄游通道,使短颌鲚等洄游性鱼类以及草鱼、青鱼等中下层植食性鱼类丧失产卵场。湖区周边道路桥梁施工如基坑开挖、材料堆放及机械作业扰动了局部底质,破坏了底栖生物栖息环境,进而影响鱼类摄食链;桥墩等构筑物改变水流形态,形成了不规则漩涡,可能干扰短颌鲚等敏感洄游鱼类的迁移路径,增加其寻找产卵场的阻力。持续干旱导致水域萎缩、水深降低,使多层次水生生境单一化,压缩了不同水层鱼类的栖息空间。菱角密集覆盖抑制了水体溶解氧交换,根系竞争破坏水生植物多样性,减少植食性鱼类食物来源;菱角机械收割会直接损伤水生植物和浮游生物,破坏产卵基质与幼鱼隐蔽场,降低幼鱼存活率,同时搅动底泥引发水体浑浊,影响鱼类捕食并释放底泥有害物质。聚类分析表明,城西湖鱼类群落的聚类距离显著增大,优势种集中于鲫鱼、䱗等耐污性强、栖息地需求低的小型鱼类,洄游性及中下层植食性鱼类的种类与数量持续减少,群落生态功能弱化、稳定性下降,凸显栖息地单一化的严重限制。
3.3.3 水体富营养化
水体TN、TP超标会促进浮游生物增殖,为浮游食性鱼类(如似鱎)提供饵料,但抑制了清水性物种。城西湖水体污染导致氮、磷负荷升高,进一步强化了杂食性鱼类的竞争优势[26]。
3.4 结论
城西湖2022—2024年水体处于富营养化状态、鱼类群落结构呈现“定居种恢复,洄游种衰退”的特征(鲫、似鱎等耐污种持续占优;短颌鲚因水文连通丧失而局部衰退),下一步可通过闸坝生态调度(在鱼类繁殖季节,通过调节闸坝放水,模拟自然的洪水过程,为鱼类创造适宜的繁殖环境,促进其产卵和幼鱼孵化帮助洄游性鱼类克服闸坝的阻隔,恢复其正常的洄游路线,加强物种间的交流,从而丰富群落物种组成,改变群落简化趋势)、磷负荷削减(加强生活污水尤其是农业面源污染的治理,严格控制含磷废水的排放)、营养级重建(根据生态系统的原有结构和功能,有针对性地引入或恢复关键物种,重塑稳定的营养级关系)三路径协同,改变群落简化趋势,重点突破闸坝阻隔与富营养化治理瓶颈。