

森林是陆地上面积较广、生物多样性较丰富的生态系统之一,不仅为陆生动植物提供了栖息地,还通过碳汇功能、气候调节、水源涵养、土壤保持等生态服务功能维持生态平衡和人类社会可持续发展[1-2]。随着农田、商用林地和城市的快速扩张,森林破坏和退化问题日益严重[3],森林的面积、稳定性、恢复力及其提供生态系统服务的能力不断下降[4],生物多样性丧失加剧[5]。自然恢复易受到土壤退化、气候变化和物种流失的影响,难以在短时间内恢复完整的森林生态系统,因此,人工主导的森林修复是森林生态恢复的重要手段[6-7]。
当前,森林修复实践多侧重于植被群落的恢复及其对生态系统结构的改善作用,但单纯依赖植被恢复并不足以全面修复森林的生态功能。动物群落在维持生态系统的稳定性和功能完整性方面发挥着重要作用,动物群落多样性丧失会对森林植被的维持和更替产生负面影响[8]。鸟类是森林食物网的重要组成部分,其取食花蜜和果实的行为能够帮助花粉和种子传播,促进植被的更新和扩展[9-10];其捕食活动发挥着植物—植食昆虫的调控作用,是控制森林害虫的重要手段[11]。健康的鸟类群落整体上增强了森林生态系统的功能与多样性,是维持植物群落动态平衡的重要环节。此外,鸟类相对易于观察,并且不同类型的森林结构能够为不同鸟类物种提供适宜的栖息环境,鸟类物种被广泛用于生物多样性的长期监测[12]。因此,在森林生态恢复过程中,关注鸟类群落的动态及其与林分特征变化的关系,优化鸟类栖息地,对提升森林生态系统的服务功能、增强森林生态恢复的可持续性具有重要作用[8]。
目前已有较多研究探讨鸟类群落对人工次生林中不同林分特征的响应。Komlós等[13]研究发现,树木高度是预测鸟类物种丰富度和群落组成的重要变量,高大的乔木层和密集的灌木层通常会增加鸟类的物种丰富度。此外,林分年龄对鸟类多样性也有重要影响。Birčák等[14]研究指出,林龄较高的林分鸟类物种丰富度较高,可能与老龄林提供更多资源(树洞、枯木等)以及复杂的垂直结构有关。然而,Abbas等[15]研究发现,在某些情况下,中龄林可能比老龄林具有更高的物种丰富度,可能是因为中龄林在生态演替过程中为物种提供了更丰富的食物资源和栖息地。人工修复次生林中的树种组成和植被结构对鸟类功能性群体的影响也存在争议。John等[16]研究表明,混交林比纯林更有利于提高鸟类多样性,因为混交林为其提供了更多样化的资源和微生境。然而,在某些情况下,纯林可能通过简化竞争环境帮助体型较小、食性专一的鸟类等群体选择栖息地[17]。此外,植被郁闭度和盖度也是影响鸟类选择栖息地的重要因素。郁闭度较高的林分可能为一些偏好密闭环境的鸟类提供庇护,但同时也可能限制阳光需求较高或飞行范围较大的物种[18-19]。Stouffer等[20]研究指出,次生林中的某些功能性群体数量(穴居鸟类或食虫鸟类)明显减少,可能与次生林中植被结构均一化以及资源可利用性下降有关。因此,人工修复次生林中不同林分特征对鸟类分布和群落结构的影响亟须进一步研究。
强化原生动植物保护、维护生态系统完整性,是自然保护地建设的核心目标之一。因此,深入研究和分析影响森林物种分布、群落多样性及其动态的关键因素,可为森林管理、保护和恢复提供理论依据和实践指导。广东省佛山市云勇林场是典型的人工修复次生林,本研究以此为研究地点,对林场内的鸟类群落进行调查,分析该林场中不同地点、不同生长类型植被的高度、盖度、生物量、群落构成等林分特征之间的差异,及其如何介导局地尺度内鸟类分布和群落构成,探讨鸟类群落变化与森林重建过程中的植被变化之间的关系,为理解不同林分特征影响鸟类栖息地的选择及未来调整森林经营管理措施、促进生态恢复提供参考。
1 材料与方法
1.1 试验地基本情况
佛山市云勇林场位于广东省佛山市西南部,地理坐标为112°38′26″—112°42′25″ E、22°41′54″—22°46′50″ N,总面积2 007.8 hm2,年均气温22.0 ℃,偶有霜冻,雨量充沛,雨季多集中在4—8月,年均降水量2 000 mm,属于亚热带气候区。该地处于丘陵地带,土壤为花岗岩发育的酸性赤红壤,土壤pH在5.5~6.0,土层养分含量丰富;场内最高峰为鸡笼山,海拔418.7 m[21]。该林场是生产经营性林场,生产松、杉等木材。2002年,该林场转型为生态公益型林场,并陆续对不同林班中的杉木纯林进行皆伐,同时引入阔叶乡土树种进行林分改造[21]。因此,该林场的森林植被由不同年龄、不同植物群落、相对均质的次生林小班组成,有利于研究局地尺度上动物群落对森林重建和恢复过程的响应。
1.2 调查方法
2021年1月至2023年4月,采用样线法和样点法调查试验地鸟类群落。针对林场生境分布特点,共设置了8条固定样线,包括曾板田(ZBT)、十二沥(SEL)、云勇村(YOC)、云益村(YYC)、鸡笼山(JLS)、车子道(CZD)、山塘(SHT)和场部(CHB),样线长度1.0~1.8 km,尽量穿过不同龄组的小班。样线上每隔200 m设1处固定样点,样点监测半径50 m,每个样点停留计数10 min。每3个月进行1次样线调查(2022年夏季未进行调查),至2023年4月结束,共调查了12次。在晴朗无风的天气开展调查,在日出至10:30前或15:30至日落前进行,调查人员记录观察到的鸟类物种、个体数量、行为等信息,每条样线调查时间2~3 h。超过监测半径50 m以及短时间内大范围活动的鸟类数据在分析时不予采用,最终乌鸦、猛禽、燕、雨燕、䴙䴘等13种鸟类不用于数据分析。
调查期间,在沿样线的13个位点上布设红外相机对在地面及林下层活动的鸟类进行补充调查,包括CZD 4台、JLS 3台、SEL 3台、YYC 2台以及ZBT 1台。相机布设在样线两侧50 m距离内的乔木或灌木下。相机设置为触发后拍摄3张照片并录制15 s视频,触发间隔为10 s。每季度进行样线调查时检查各红外相机,回收数据并更换电池。
1.3 测定项目及方法
1.3.1 鸟类群落构成
由于样线、样点法和红外相机法对物种个体数量估算的假设不同,且红外相机法更偏向对森林下层和地面活动鸟类的监测。因此,后续分析中分别基于观察数据和红外相机数据对鸟类群落进行独立的计算与比较。基于观察数据的鸟类群落构成,使用各样线上记录到的物种及其个体数量衡量;基于红外相机数据的鸟类群落构成,通过每100 d拍摄到的独立有效照片数量计算得到的目标物种相对密度(Relative abundance index,RAI)来衡量[22],RAI计算如式(1) 。
RAI =
式中,Ni 为相机位点i拍摄的某一物种的有效照片数;CameraDay i 为相机位点i的拍摄天数,其根据各个样线利用红外相机拍摄的总天数计算,其中CZD 2 702 d,JLS 1 950 d,SEL 2 079 d,YYC 1 290 d,ZBT 731 d。
1.3.2 不同样线鸟类群落差异
林分特征提取自2022年佛山市森林资源调查数据中的云勇林场小班数据。林分特征有草本层、灌木层、下木层和乔木层4个生长型,其中各层数据均包括平均高度(cm)、盖度(%)和生物量(kg/hm2),灌木层、下木层还包括平均地径(cm)、树龄(a)和株数(株/hm2),乔木层还包括平均胸径(cm)、树龄、株数和郁闭度(%)。小班数据被转化为分辨率为10 m的栅格,使用上述数据的平均值以及森林群落结构、树种结构和森林健康度的众数表示样线两侧50 m以内各生长型林分特征的整体情况,使用上述各林分特征的标准差表示林分特征的变异程度。
使用生物—环境匹配分析和冗余分析计算不同样线的鸟类群落及鸟类物种对林分特征的响应。生物—环境匹配分析可以筛选出最优的林分特征子集,使得不同样线之间的林分特征差异与鸟类群落相异性之间的相关性最大。通过Bray-Curtis相异度指数计算各样线鸟类群落的相异性,同时通过聚类分析判断鸟类群落构成相似的样线。使用最大—最小值法将草本层、灌木层、下木层和乔木层各特征数据(除森林群落结构、树种结构和健康度)标准化为0~1的数据;用Gower距离计算不同样线间的林分特征差异;使用Pearson相关系数衡量鸟类群落相异性和林分特征差异之间的相关性。为节约计算时间,林分特征子集设置为最多可包含7个变量。通过生物—环境匹配分析获得的林分特征子集被用于冗余分析,对鸟类群落进行约束性排序。冗余分析第1和2轴的解释力,以及各林分特征和鸟类物种的得分,用来推断不同物种对林分特征的响应。
1.3.3 物种分布模拟
将观察数据和红外相机数据合并后建立物种分布模型,合并后仅有21种鸟类的记录位点超过10个。因此,将林分特征作为环境图层,使用最大熵(MaxEnt)模型预测各鸟类物种在林场的潜在分布,并根据其对模型的贡献判断对各物种分布影响较大的林分特征。首先,建立在样线周边50 m范围内的各物种分布模型,然后将模型投影到有全部测量数据的小班中。模型模拟的鸟类至少有3个相互距离超过50 m的观测记录,每次随机生成300个伪无分布点;每次建模时将70%的记录作为训练集,30%的记录作为验证集,其余使用软件的自动设置;使用受试者特征曲线(Receiver operating characteristic curve,ROC)以及曲线下方面积(Area under curve,AUC)作为判断模型优劣的标准,AUC值接近1,表明模型对物种分布有较好的预测水平。
1.4 数据分析
不同样线鸟类群落差异比较中,使用betapart包计算Bray-Curtis指数,使用vegan包进行其余分析;物种分布模拟中,使用MaxEnt(Ver. 3.3.3.3)软件[23]建立模型。
2 结果与分析
2.1 鸟类多样性
由表1可知,2021年1月至2023年4月,通过样线法和红外相机法,在8条样线中共记录到103种鸟类,其中样线法调查到87种,共1 806个个体;红外相机法调查到32种。ZBT、SEL、YYC、JLS和CZD的RAI最高的物种分别为棕颈钩嘴鹛(Pomatorhinus rufieollis)(0.68)、黑冠鳽(Gorsachius melanolophus)(1.68)、红胁蓝尾鸲(Tarsiger cyanurus)(0.39)、灰背鸫(Turdus hortulorum)(0.72)和虎斑地鸫(Zoothera dauma)(1.15)。样线法在ZBT和CHB记录到的物种数较多,可能与这两处地点周边生境类型较为丰富有关。ZBT和CHB周边次生林小班林龄长、林下植被丰富,同时被小块农田、菜园、花圃、村舍等人工景观环绕,能够为不同生态位的鸟类提供休憩和觅食场所。红外相机法在ZBT只记录到4种鸟类,可能是由于此处仅布置了1台红外相机,总拍摄天数较短;SEL和CZD则记录了较多的林下活动鸟类。
表1 试验点的各样线鸟类物种数 |
| 样线 | 物种数 | |
|---|---|---|
| 样线法 | 红外相机法 | |
| ZBT | 52 | 4 |
| SEL | 29 | 16 |
| YOC | 36 | - |
| YYC | 36 | 14 |
| JLS | 25 | 10 |
| CZD | 30 | 20 |
| SHT | 37 | - |
| CHB | 54 | - |
| 总计 | 87 | 32 |
|
2.2 鸟类群落差异
2.2.1 Bray-Curtis指数分析
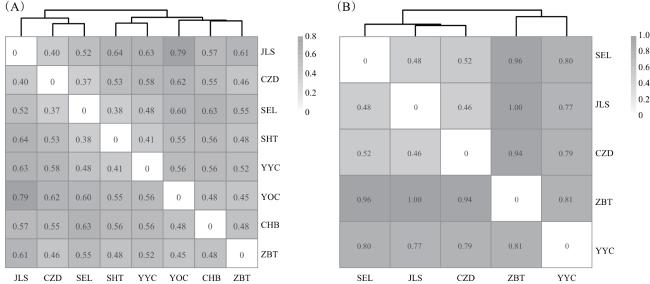
由图1可知,基于观察数据和红外相机数据计算的多位点Bray-Curtis指数分别为0.748和0.794,表明在考虑相对密度的情况下,试验地样线间鸟类群落差异较大。聚类结果表明,根据群落构成,观察数据可将8条样线聚类为3组,其中JLS、CZD和SEL聚类为一组,SHT和YYC为一组,CHB、YOC和ZBT为一组。红外相机数据同样将JLS、CZD和SEL聚类为一组,说明这3条样线的鸟类群落较为相似。生物—环境匹配分析结果表明,草本盖度均值、灌木高度均值以及下木生物量的标准差是解释观察数据鸟类群落差异的最佳林分特征组合(r=0.492);下木生物量均值、乔木年龄均值和乔木生物量均值是解释红外相机数据的最佳林分特征组合(r=0.842)。
2.2.2 冗余分析
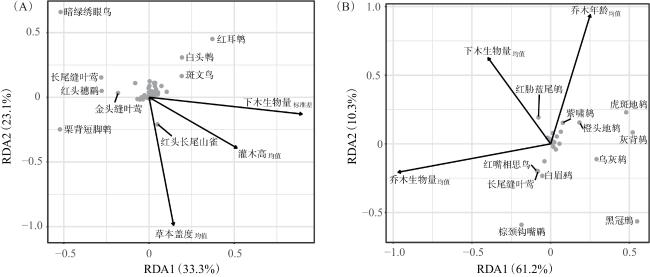
冗余分析结果表明,大部分物种的排序集中在原点,只有少数物种显示出明显的林分特征偏好。由图2A可知,约束性部分可以解释样线法数据56.4%的方差,其中前两轴分别可以解释33.3%和23.1%的方差。下木生物量标准差和灌木高度均值对轴1的解释力较强,其与该轴呈正相关,红耳鹎(Pycnonotus jocosus)、白头鹎(Pycnonotus sinensis)和斑文鸟(Lonchura punctulata)等在该轴正向得分较高,暗绿绣眼鸟(Zosterops japonica)、栗背短脚鹎(Hemixos castanonotus)、长尾缝叶莺(Orthotomus sutorius)、红头穗鹛(Stachyris ruficeps)和金头缝叶莺(Orthotomus cucullatus)则在该轴负向得分较高 。草本盖度均值对轴2的解释力较强,其与该轴呈负相关,暗绿绣眼鸟、红耳鹎、白头鹎、斑文鸟和长尾缝叶莺在该轴正向得分较高,栗背短脚鹎和红头长尾山雀(Aegithalos concinnus)在负向得分较高。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
由图2B可知,约束性部分可以解释红外相机数据71.5%的方差,前两轴分别可以解释61.2%和10.3%的方差。乔木生物量均值和下木生物量均值对轴1的解释力较强,且二者与轴1呈负相关,黑冠鳽、灰背鸫、虎斑地鸫、乌灰鸫(Turdus cardis)和橙头地鸫(Zoothera citrina)在该轴正向得分较高,棕颈钩嘴鹛在该轴负向得分较高;乔木年龄均值和下木生物量均值对轴2的解释力较强,二者与轴2呈正相关,虎斑地鸫和红胁蓝尾鸲在该轴正向得分较高,棕颈钩嘴鹛、黑冠鳽、白眉鹀(Emberiza tristrami)、红嘴相思鸟(Leiothrix lutes)和长尾缝叶莺在该轴负向得分较高。
2.3 鸟类分布
本研究为林场内的58种鸟类建立了MaxEnt分布模型,其中11种鸟类的AUC超过0.7,28种鸟类的AUC在0.5~0.7。通过对41种鸟类分布模型中不同林分特征的贡献率分析发现,在人工重建的小班内,与植被的高度、年龄、生物量等因素相比,森林群落结构和树种结构对预测鸟类分布的贡献更大。在AUC>0.5的39种鸟类模型中,树种结构、森林群落结构、森林健康度、下木地径、草本高度、乔木高度、灌木盖度分别在22、14、13、10、7、4和1种鸟类模型里的贡献度超过20%。
3 结论与讨论
立地构建初期,乔木层植株低矮,灌木层发育不充分,草本层易被少数喜阳优势物种占据;此时森林垂直结构缺失,林下微气候昼夜变化明显,仅先锋物种具备生境适应性。随着乔木层持续生长、灌木层充分发育,草本层受遮阴、动物采食等因素影响,水平异质性显著提升;在此过程中,森林群落结构日趋复杂,林下微气候逐渐稳定,生态系统生产力逐步提高,更多动植物找到适配的微生境,森林生物多样性进入稳步上升阶段。随着林冠盖度进一步增长,林下光照变得微弱,原有的灌木层和草本层逐渐被耐阴、矮小的植物替代,中下层植被趋向简单化,植物群落的多样性和垂直结构的复杂性均有所缺失[26]。在演替后期,昆虫、鸟类、兽类依赖植被提供食物和栖息地,其多样性通常与林冠郁闭度增加、植被垂直结构复杂性降低呈钟形或负相关[27]。
本研究发现,不同试验点的鸟类偏向选择有不同林分特征的生境。红耳鹎和白头鹎偏向下木更茂密、灌木层更高的生境,而暗绿绣眼鸟、栗背短脚鹎则偏向下木层相对稀疏、灌木层较矮的生境;虎斑地鸫、灰背鸫和黑冠鳽等均偏好林龄更长、乔木相对稀疏的生境,但虎斑地鸫更偏好下木茂密的林地。不同鸟类对树种结构也表现出不同的偏好,在MaxEnt模型中,树种结构对红耳鹎、棕颈钩嘴鹛和画眉(Garrulax canorus)的生境偏好贡献率均超过40%,其中红耳鹎偏好阔叶纯林和阔叶相对纯林,棕颈钩嘴鹛偏好针叶混交林,画眉则偏好阔叶相对纯林和针叶纯林。分布模型中,鸟类对森林植被的群落结构也表现出不同偏好,例如,白喉红臀鹎(Pycnonotus aurigaster)和珠颈斑鸠(Streptopelia chinensis)偏好结构完整的植被,在树林中层和下层活动的红嘴相思鸟、金头缝叶莺等则偏向结构较完整的植被,常在林中层和冠层活动的赤红山椒鸟(Pericrocotus flammeus)和栗背短脚鹎则偏向简单结构的植被。以上结果表明,在森林恢复过程中,需根据物种的栖息需求调整管理措施,同时还要维持微生境的多样性以满足不同物种的环境需求。具体而言,同样是需要林下环境的物种,黑冠鳽可通过选择性修剪林下植被,营造相对开阔的林下环境,为其提供适宜的栖息条件;而对虎斑地鸫则要保持较高的灌木层和下木层。对于喜好开放环境的物种,则应加强林地的开放性设计,适度开展疏伐、间伐等经营抚育措施,避免过度郁闭的林地环境。
综上,森林恢复不仅是植被恢复的过程,还应考虑不同物种的栖息需求。在设计和开展森林经营管理措施时需考虑对野生动物,特别是对鸟类等关键物种栖息地的优化。通过设计合适的栖息环境,提升森林的生物多样性和生态系统服务功能,从而实现生态系统的全面恢复。