

赤芍(Paeonia lactiflora)是芍药科芍药属植物,含有芍药苷等成分,具有清热凉血、散瘀止痛等功效。该植物在我国栽培范围较广泛,山东菏泽、河南洛阳等地是较为出名的产地,近年在东北等地也有引种。赤芍种子具有上下胚轴双重休眠的特性[1]。陶新宇等[2]研究发现,通过机械破除种皮的方式,可提高芍药种皮通透性和发芽率。于成波[3]研究发现,低温层积60 d且使用浓度为20×10-6的细胞分裂素能缩短下胚轴萌发时间,提高生根率和发芽率。秦立金等[4]研究表明,60 ℃干热处理有利于打破赤芍种子休眠。相关研究表明,赤霉素(GA3)对打破植物种子休眠、促进中胚后熟的效果显著,但目前关于赤芍种子上下胚轴休眠的研究有待进一步深入。越冬低温是东北地区引种赤芍时面临的问题之一。低温在一定程度上能够促进种子萌发,但幼苗遭受低温易形成冻害。施加外源GA3等植物激素有助于植物幼苗抵御低温胁迫[5-6],但对于其在幼苗种子萌发期施加是否能够促进幼苗的耐低温性暂未有较多研究。本研究选取3个不同生态区的赤芍种质资源,考察外源GA3和低温对赤芍种子萌发和幼苗生长的影响,为赤芍种子规模化繁殖提供参考。
1 材料与方法
1.1 试验材料
本研究收集吉林(JL)、山东菏泽(SD)、河南洛阳(HN)3个产地的赤芍作为试验材料。JL赤芍为SD赤芍引种驯化后的繁育种源。
1.2 试验设计
挑选饱满健康种子,清洗干净后在清水中浸泡48 h,吸水后沥干水分,分别放入清水(CK)、100和300 mmol/L GA3中浸泡5 d后,常温培养3 d,将CK、300 mmol/L GA3处理组的50%种子置于4 ℃下处理4 h,其余种子继续常温处理,将种子播种于装有蛭石的育苗盘中,每组100粒种子,试验重复3次。萌发试验使用人工气候箱(ZPW-400),光照6级/光照0级,光周期12 L∶12 D,湿度80%,白天、晚上温度分别为28、25 ℃,统计各处理组90 d后生根率、根长等指标。清水处理后的种子常温萌发且下胚轴长度>15 cm时,喷施清水(CK)、100和300 mmol/L GA3,将育苗盘在4 ℃环境中放置4 h,检测幼苗的丙二醛(MDA)含量、过氧化氢酶(CAT)和过氧化物酶(POD)活性,每个处理10株幼苗,试验重复3次。
1.3 试验项目及方法
1.3.1 种子发芽率测定
生根率(%)=萌发种子数/种子总数×100
发芽率(%)=发芽生苗种子数/种子总数×100
发芽势(%)=达到高峰日时的发芽数/种子总数×100
1.3.2 抗氧化酶活性测定
称量0.5 g赤芍胚根,加入适量100 mmol/L磷酸缓冲液(pH 6.0)研磨至匀浆,4 ℃离心5 min,取上清液定容至100 mL。在对照比色皿中加入3 mL反应液(体积分数为0.065的愈创木酚、体积分数为0.038的30% H2O2溶于100 mmol/L磷酸缓冲液)和1 mL磷酸缓冲液。样品比色皿中加入3 mL反应液和1 mL酶提取液,在470 nm波长下检测吸光度,每隔1 min读数1次。
POD活性[U/(g·min)]=ΔA470×VT /m×Vs ×0.01×t
式(4)中,ΔA470表示反应时间内470 nm波长下吸光度变化值;VT 表示提取液总体积;m表示样品鲜重;Vs 表示反应提取液体积;t表示反应时间。
称量0.5 g赤芍胚根加入适量0.2 mol/L磷酸缓冲液(pH 7.8,加入1%聚乙烯吡咯烷酮)研磨至匀浆,转入25 mL容量瓶中定容,于5 ℃冰箱静置10 min。取上清液(反应液)4 000 r/min离心5 min。对照组比色皿中加入0.2 mL酶液(灭活)、1.5 mL磷酸缓冲液(pH 7.8)、1.0 mL蒸馏水和0.3 mL 0.1 mol/LH2O2,在240 nm波长下检测吸光度A0,每隔1 min读数1次,连续测4次。样品比色皿加入有活性酶液,重复2次,检测吸光度A1、A2。吸光度值与CAT活性计算如式(5 )~(6 )。
ΔA240=A0-(A1+A2)/2
CAT活性[U/(g·min)]=ΔA240×VT /m×Vs ×0.1×t
式中,ΔA240表示反应时间内240 nm波长下吸光度变化值。
1.3.3 可溶性糖和可溶性蛋白含量测定
称量新鲜胚根0.5 g,加入装有10 mL蒸馏水试管中沸水浴1 h,将提取液定容至25 mL。吸取1 mL提取液,加入1 mL蒸馏水、0.5 mL蒽酮乙酸乙酯和5 mL浓硫酸,充分震荡,沸水浴1 min后冷却至室温,630 nm波长下测吸光度。以不同浓度纯蔗糖制作标准曲线,计算可溶性糖含量。称取0.5 g胚根冻干粉末,加入磷酸缓冲液研磨成匀浆后,离心10 min,取上清液为提取液。吸取1.0 mL提取液,加入5 mL考马斯亮蓝试剂,摇匀,放置2 min后在595 nm波长下测量吸光度。将不同浓度的牛血清白蛋白溶液显色并测定吸光度,制作标准曲线,以外标法计算可溶性蛋白含量。
1.3.4 丙二醛MDA含量测定
称取0.5 g胚根样品,加入5 mL磷酸缓冲液冰浴研磨成匀浆,离心后取上清液为提取液。取2 mL提取液,加入体积分数为0.67%硫代巴比妥酸溶液2 mL,沸水浴15 min,冷却后离心取上清液检测660、532和450 nm波长下的吸光度。对照组提取液为蒸馏水。
MDA含量[μmol/g·FW]=[6.45×(ΔA532-ΔA600)-0.56×ΔA450]×VT /FW×Vs
1.4 数据分析
使用SPSS 17.0软件进行单因素方差分析(One-way ANOVA),用Duncan’s检验进行统计学分析,使用Microsoft Excel软件作图。
2 结果与分析
2.1 对不同种源赤芍种子生根的影响
试验观察发现,常温下下胚轴先于上胚轴萌动。由图1可知,外源GA3浓度越高,其促进赤芍生根作用越明显。CK处理的JL、SD和HN芍药种子的生根率、根长差异无统计学意义(P>0.05),100和300 mmol/L GA3处理下JL芍药种子的生根率差异无统计学意义(P>0.05)。4 ℃低温使JL、SD和HN芍药种子生根率和根长下降;4 ℃+300 mmol/L GA3处理下的生根率较4 ℃处理组提高了355.28%、296.44%和130.81%,根长提高了160.34%、190.85%和93.25%。
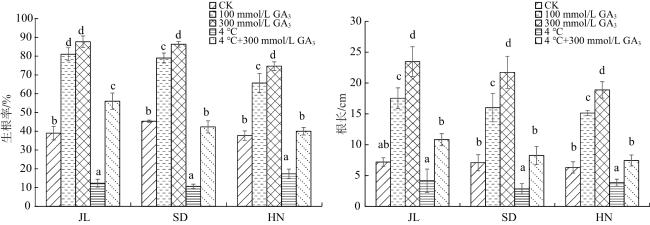
2.2 对不同种源赤芍种子发芽的影响
由表1可知,不同种源的CK发芽时间差异不明显,均在170 d以上。低温会延长发芽时间,抑制赤芍种子发芽,使JL、SD和HN赤芍种子发芽率分别降低2.67、5.33和3.67个百分点。外源GA3处理会促进种子生芽,300 mmol/L GA3的发芽率较CK明显提高,发芽势较CK明显增强,且差异具有统计学意义(P<0.05);GA3浓度越高,促进发芽作用越明显。
表1 低温和外源GA3对赤芍种子发芽的影响 |
| 种源 | 处理 | 发芽天数/d | 发芽率/% | 发芽势/% |
|---|---|---|---|---|
| JL | CK | 174.00±13.75 c | 9.00±3.00 ab | 7.67±1.53 b |
| 100 mmol/L GA3 | 138.00±16.70 ab | 18.00±2.65 c | 11.67±1.42 c | |
| 300 mmol/L GA3 | 121.33±11.50 a | 28.33±2.08 d | 17.67±2.52 d | |
| 4 ℃ | 198.00±7.11 d | 6.33±0.96 a | 3.33±0.22 a | |
| 4 ℃+300 mmol/L GA3 | 154.00±9.26 bc | 12.00±1.22 b | 6.00±0.86 b | |
| SD | CK | 172.33±14.29 bc | 10.00±1.73 b | 6.10±1.48 b |
| 100 mmol/L GA3 | 138.33±4.51 ab | 17.67±1.53 e | 9.14±1.32 c | |
| 300 mmol/L GA3 | 120.00±4.36 a | 27.67±3.06 d | 17.56±1.48 d | |
| 4 ℃ | 201.00±8.39 c | 4.67±0.38 a | 2.00±0.58 a | |
| 4 ℃+300 mmol/L GA3 | 166.00±9.97 bc | 13.67±1.43 c | 5.00±1.00 b | |
| HN | CK | 175.33±14.50 cd | 11.00±2.65 ab | 3.94±1.57 ab |
| 100 mmol/L GA3 | 146.00±12.49 b | 17.33±1.15 b | 9.11±0.28 c | |
| 300 mmol/L GA3 | 117.33±6.66 a | 26.33±4.51 c | 16.63±2.01 d | |
| 4 ℃ | 190.00±13.53 d | 7.33±2.31 a | 2.00±0.57 a | |
| 4 ℃+300 mmol/L GA3 | 164.33±8.33 c | 11.00±2.65 ab | 4.00±0.58 b |
|
2.3 对不同种源赤芍幼苗抗氧化能力的影响
2.3.1 抗氧化酶活性
由图2可知,在常温下,外源GA3处理的JL赤芍幼苗POD活性明显高于CK,差异具有统计学意义(P<0.05)。低温处理下3个产地赤芍的POD活性较常温明显提高,说明赤芍幼苗中的POD也可能参与抵御低温的环节。
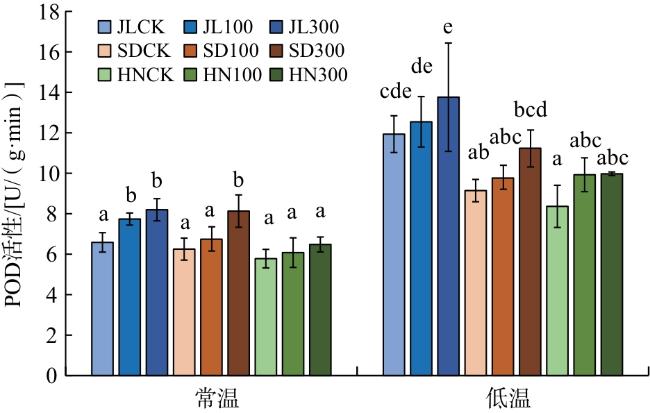
由图3可知,300 mmol/L GA3处理后3个产地赤芍幼苗CAT活性较CK明显提高,差异具有统计学意义(P<0.05)。与常温相比,低温条件下3个产地赤芍的CAT活性明显上升;相同处理下,JL赤芍的CAT活性明显高于其他2种赤芍。综上,说明JL赤芍经过多年引种驯化提升了低温下CAT活性,进而更加适应低温环境。
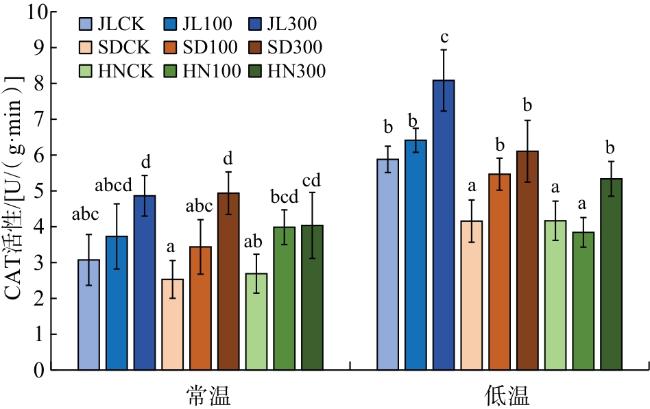
2.3.2 可溶性糖和可溶性蛋白含量
由图4可知,常温下,300 mmol/L外源GA3处理的赤芍幼苗可溶性糖含量明显高于CK,其中JL、HN赤芍差异具有统计学意义(P<0.05)。低温下,JL赤芍幼苗的可溶性糖含量明显高于其他2个产地赤芍。4 ℃+300 mmol/L GA3处理下,JL赤芍幼苗的可溶性糖含量较SD、HN赤芍分别提高13.90%、18.83%。
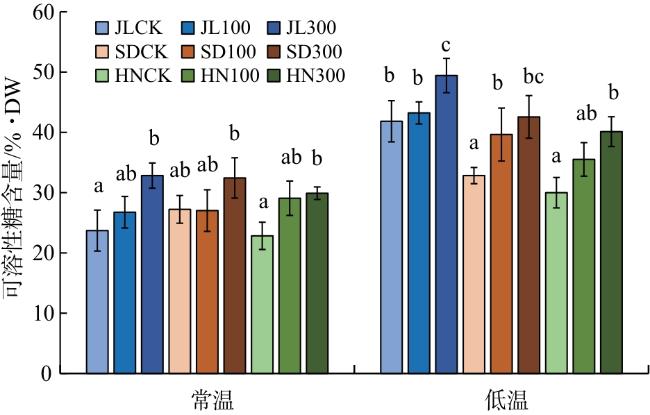
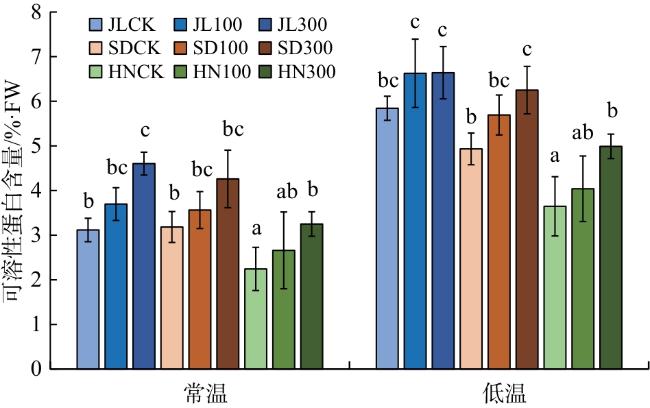
由图5可知,常温下,300 mmol/L外源GA3处理后的JL、HN赤芍可溶性蛋白含量较CK分别增加47.44%、45.09%,差异具有统计学意义(P<0.05)。与常温相比,低温处理下3个产地赤芍可溶性蛋白含量明显上升,其中以4 ℃+300 mmol/L GA3 处理下的JL赤芍的可溶性蛋白含量最高,较SD和HN增加16.24%和33.07%。
2.3.3 MDA含量
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
外源GA3会加速细胞分裂和细胞伸长,对赤芍、凤丹等芍药科植物种子萌发有一定的促进作用[7]。本试验结果表明,赤芍种子萌发时上下胚轴打破休眠时间存在差异,其中发芽需要更多时间;外源GA3可促进3个产地赤芍种子生根和出芽,缩短胚根和胚芽萌动的时间,提高发芽率和发芽势,以300 mmol/L GA3的促进作用最明显。
可溶性糖和可溶性蛋白是植物细胞中重要的渗透调节物质,细胞遭受低温伤害时渗透调节物质能提高细胞内溶质浓度,保护质膜,从而增强植物抗胁迫能力[10]。常温下,3个产地赤芍可溶性糖含量无明显差异,外源GA3在一定程度上能增加可溶性糖和可溶性蛋白含量,以300 mmol/L GA3的促进较好。低温会提高赤芍幼苗的可溶性糖和可溶性蛋白含量,以JL赤芍的可溶性糖和可溶性蛋白含量上升幅度最大,其可能通过维持渗透调节物质含量提高抗低温能力。
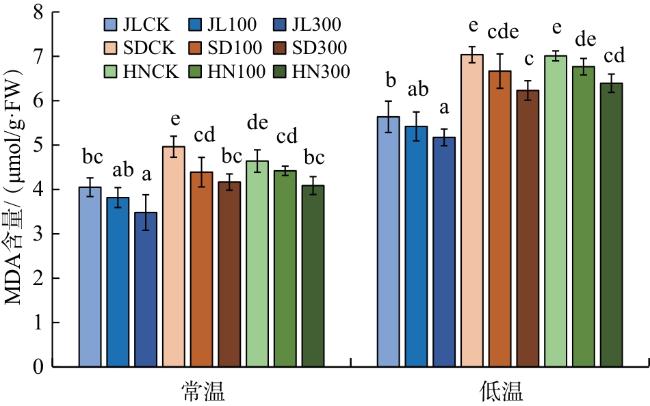
越冬能力是赤芍在东北地区栽培的关键限制因素。低温下,MDA含量能够直观反映植物细胞受伤害的程度,抗低温能力强的种质资源MDA含量上升幅度更小[11]。本研究中,与常温相比,低温处理后3个产地赤芍的MDA含量均有所上升,以JL赤芍MDA含量最低,较常温处理上升幅度最小,说明低温对于SD、HN赤芍胚根细胞的破坏作用比JL赤芍更显著;300 mmol/L外源GA3抑制MDA产生的效果较好,说明外源GA3能够提高赤芍抗低温的能力。
综上,外源GA3有利于促进赤芍萌发和发芽,一定程度上能增强赤芍抗低温能力,其中JL赤芍经过多年引种驯化更加适应研究区的低温环境,受到低温伤害的概率更小。