

在自然条件下,干旱往往伴随着土壤盐渍化,盐胁迫是对植物损害较为严重的非生物胁迫之一,会引起植物生长、形态结构和生理生态等一系列过程变化,影响植物的生长发育,甚至引起植株死亡。生长在盐渍化地区的植物,为适应环境进化出各自特有的耐盐机制。目前,通过选育耐盐碱作物品种及开发相关植物资源,成为提升盐渍化土地利用效率的重要策略[1]。
盐胁迫对植物的不利影响主要涉及离子和渗透物质因素,二者均会导致植株无法吸收水分,最终造成植物生长速率降低,直至死亡。大多数植物对盐较敏感,也有少部分物种具有耐盐性,其主要通过减少Na+吸收、将Na+排出、区隔化等途径来减轻盐胁迫对自身的伤害[2]。虽然Na+对大多数植物具有毒害作用,但对于少数植物,如C4植物和盐生植物,Na+的缺乏可能使其无法完成生活史[3]。K+对植物的生长发育、生理代谢及抗逆性等具有多方面的影响。然而,K+与Na+存在显著的离子拮抗作用,因此盐胁迫常导致植物体内K+亏缺,而耐盐物种可通过根系的K+选择性吸收及转运系统优先维持K+稳定,从而提高K+/Na+比来增强其耐盐性[4]。可见,保持K+稳态平衡是提高植物耐盐性的重要策略[5]。此外,当植物受到高盐胁迫时,一方面会引起光合作用效率下降致使其生长发育受到抑制;另一方面,K+参与光合作用中碳水化合物的合成和转运,而Na+的大量积累会导致K+的亏缺,由此进一步影响植株的光合作用[6-7]。
在盐渍化土壤环境中,可溶性盐类可解离出Na⁺、Cl⁻等离子形态。当植物根系通过质外体途径进行水分运输时,这些溶解态离子会伴随蒸腾流发生共质体—质外体双向迁移,导致叶片中积累大量Na+。然而,高浓度的Na+对耐盐植物造成的损害较小,此类植物对细胞中的Na+有较强的区域化能力,即将进入细胞中的Na+积累至液泡中,从而避免因离子在细胞质等部分重要细胞器中过量积累对植株造成严重损害[6]。黑果枸杞(Lycium ruthenicum)为茄科枸杞属灌木,具有较高的药用、经济、饲用及生态价值,是荒漠地区特有的野生植物[8-9]。黑果枸杞具有很强的耐盐碱性,其在土壤全盐量达12.16%的生境仍能生长[10];如在青海诺木洪盐化荒漠区分布有大面积的黑果枸杞灌丛,且在土壤60 cm土层全盐含量低于6%的条件下,黑果枸杞长势良好[9,11]。这一现象说明黑果枸杞可能具有将Na+吸收并区隔化到叶片液泡中的功能。因此,有必要进一步探讨其在盐胁迫下的生理响应,为培育耐盐品种提供依据。本研究拟分析不同浓度NaCl处理对黑果枸杞生长发育、Na+、 K+含量及光合能力的影响,为阐明黑果枸杞对盐胁迫的适应策略提供参考。
1 材料与方法
1.1 材料培养
种子来源:黑果枸杞种子于2022年8月采自甘肃省民勤治沙综合试验站(38°34´ N,102°58´ E)。
试剂:NaCl(分析纯),Hoagland营养液,购自上海生工生物工程技术服务有限公司。试验仪器:2655-00型火焰光度计(Cole-Parmer Instrument Co.),冰点渗透压计(OSMOMAT 3000),LI-6400便携式光合仪(LI-COR Biosciences)。
1.2 发芽试验
黑果枸杞种子清洗干净后用蒸馏水浸泡12 h左右,将种子置于发芽袋(长×宽为35 cm×32 cm)中进行萌发。将Hoagland营养液分别配成含0(CK)、50、100、200、300和400 mmol/L NaCl的培养液,取20 mL分别缓慢加入袋中至完全浸湿状态,将种子置于由纸芯形成的凹槽中,袋子垂直放置在培养架上,以便直观地观察根系生长状况。每2 d更换1次营养液,以维持营养液中NaCl浓度稳定。每个处理设3次重复,每个重复40粒种子。每天观察并记录发芽情况,21 d结束发芽试验,并测量子叶直径、胚轴及初根长度。培养箱温度设为白天(28±2)℃,夜间(23±2)℃;光照时间设为16 h/d;光强度约600 µmol/(m2·s),相对湿度65%。
1.3 测定指标与方法
1.3.1 材料培养
黑果枸杞种子清洗干净后用蒸馏水浸泡12 h左右,弃去漂浮种子,将饱满种子点在铺有滤纸的培养皿中,培养室培养10 d待芽长长至1 cm左右时,挑选健壮的幼芽移栽至含有蛭石的穴盘(5 cm3,2株/盘)中,浇灌Hoagland营养液进行培养。培养室的条件设置如1.2。
1.3.2 生长指标测定
幼苗培养4~5周,进一步通过表型均一化挑选,确保幼苗大小一致,并用含0、50、100、200、300和400 mmol/L NaCl的Hoagland营养液处理幼苗21 d,进行生物量及相对生长速率测定:轻轻沿边缘剪开穴盘,收集根系;用蒸馏水冲洗植株并用吸水纸吸干水分,将各组织(根、茎、叶)迅速分离,逐一称鲜重(Wf );随后在105 ℃下杀青10 min,80 ℃烘至恒重后称干重(Wd )。相对生长速率(RGR)计算方法:RGR=(lnWf -lnWd )/△t,△t为处理天数[5]。
1.3.3 生理指标测定
(1)Na+、K+含量测定。取样过程同1.3.2所述,收集各处理的根系,根系需用预冷的20 mmol/L CaCl2浸泡2次,每次8 min,以置换细胞壁中的Na+;将烘干的黑果枸杞组织样品称重并捣碎后放入试管,加入10 mL浓度100 mmol/L的冰乙酸,并将试管用保鲜膜密封,90 ℃水浴2 h、冷却并过滤,稀释合适的倍数后,采用火焰分光光度计测定Na+、K+含量。
(2)渗透势及光合指标测定。在各浓度NaCl处理中选取发育阶段相同(叶位一致)的叶片0.5 g,立即用预冷(4 ℃)去离子水冲洗叶片表面3次,滤纸吸干表面水分,将样本投入液氮中速冻15 min,取出后塞入注射器内,温育20 min后将汁液迅速挤出,在室温条件下4 000 r/min离心1 min,取上清液50 μL,用冰点渗透压计测ic值。按照公式Ψs=-icRT计算渗透势,式中ic为上述方法测得的值,R为气体常数0.008 314,T为297 ℃[12]。
利用LI-6400便携式光合仪测定植株的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr);水分利用效率(WUE)=Pn/Tr。以上每个处理均取6个生物学重复,每个重复对应独立的植株个体。为使处理液浓度保持相对恒定,以上各处理每2 d更换一次处理液。
1.4 数据处理
试验数据统计分析采用SPSS 17.0软件,不同处理间各项指标差异采用单因素方差分析,差异显著性采用Duncan检验进行分析。用CorelDraw 12、Excel软件制图。
2 结果与分析
2.1 NaCl理对黑果枸杞种子发芽及初生苗的影响
0、50、100和200 mmol/L NaCl处理21 d的黑果枸杞的发芽率分别为35.0%、40.8%、26.7%和2.5%(图1B),表明50 mmol/L NaCl可明显促进黑果枸杞种子萌发(P<0.05)。而300和400 mmol/L NaCl处理下种子均未萌发(不再做分析)。
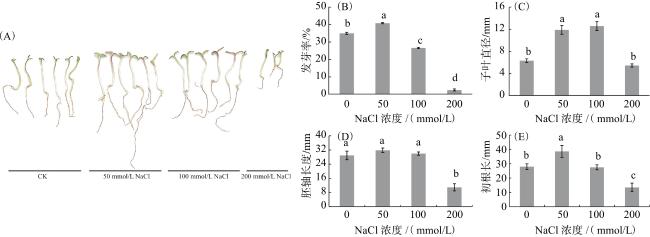
如图1A所示,黑果枸杞的初生苗在50和100 mmol/L NaCl处理下的长势优于对照(CK)。与CK相比,50和100 mmol/L NaCl处理的黑果枸杞子叶直径显著增加了87.9%、98.7%(P<0.05),而在200 mmol/L NaCl处理下下降了13.7%(图1C);与CK相比,50和100 mmol/L NaCl处理的黑果枸杞胚轴长差异无统计学意义(P>0.05),200 mmol/L NaCl处理显著下降了62.3%(图1D);与CK相比,50 mmol/L NaCl处理显著增加了黑果枸杞幼苗的初生根长(P<0.05),100 mmol/L NaCl处理的初生根长无明显变化(P>0.05),200 mmol/L NaCl处理的初生根长显著缩短了51.8%(P<0.05)(图1E)。
2.2 NaCl处理对黑果枸杞幼苗生长的影响
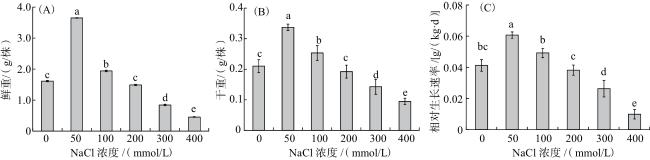
NaCl处理21 d的黑果枸杞幼苗生长情况如图2所示。与CK相比,50~100 mmol/L NaCl处理下黑果枸杞植株鲜重、干重、相对生长速率均显著提高,其中50 mmol/L NaCl处理的效果最显著,黑果枸杞鲜重增加188%,干重增加60%,相对生长速率提高49%(P<0.05)。
2.3 NaCl处理对黑果枸杞组织中K+、Na+浓度的影响
如图3所示,正常条件下(CK),黑果枸杞各组织中Na+的浓度无明显差异,50~300 mmol/L NaCl处理下,叶中Na+的浓度显著高于根和茎(P<0.05);与CK相比,50~400 mmol/L NaCl处理下黑果枸杞各组织中Na+的浓度明显增加(P<0.05),其中200 mmol/L NaCl处理下叶中Na+的浓度达到最大值,根和茎中Na+浓度在400 mmol/L NaCl下达到最大值。
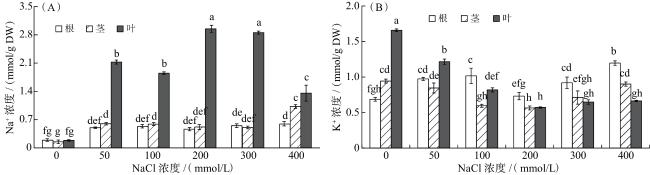
0和50 mmol/L NaCl处理下,K+主要分布在叶中,而100~400 mmol/L NaCl处理下,K+主要分布在根中。与CK相比,添加NaCl显著降低了叶中的K+浓度(P<0.05),200 mmol/L NaCl处理下达到最低,而后保持稳定;根中的K+浓度在50和100 mmol/L NaCl处理下明显增加(P<0.05),200 mmol/L NaCl处理下降低,300和400 mmol/L NaCl处理下又明显提高(P<0.05);与CK相比,100~300 mmol/L NaCl处理下茎中K+浓度显著下降,50和400 mmol/L NaCl处理下保持稳定。
2.4 NaCl处理对黑果枸杞光合指标的影响
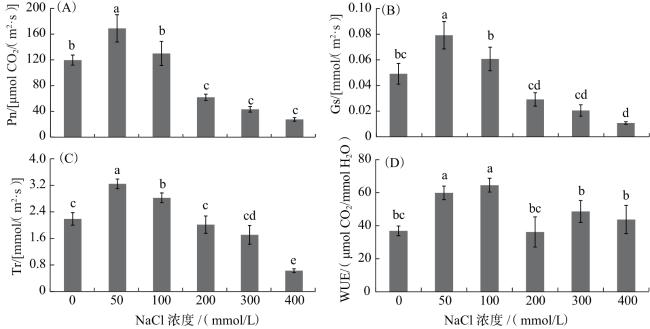
由图4可知,与CK相比,50、100 mmol/L NaCl的加入显著提高了黑果枸杞植株的Pn、Gs、Tr及WUE(P<0.05);而添加200、300和400 mmol/L NaCl植株的Pn、Gs和Tr呈现明显降低趋势。
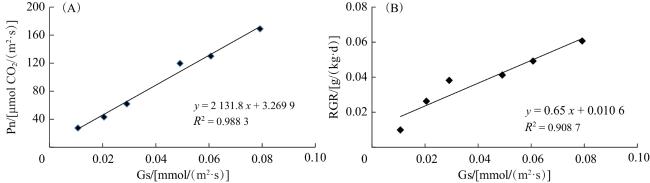
进一步分别对气孔导度与净光合速率、相对生长速率的相关性进行分析(图5),得出气孔导度与二者均存在极显著正相关关系。
2.5 NaCl处理对植株渗透势的影响
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
3.1 NaCl可通过提高Na+、K+浓度促进黑果枸杞生长
盐胁迫会抑制盐敏感植物的生长发育,并降低其光合作用和呼吸作用[3]。黑果枸杞可以在高盐渍环境下正常生长,或与柽柳(Tamarix)、盐节木(Halocnemum strobilaceum)、骆驼刺(Alhagi sparsifolia)和盐爪爪(Kalidium foliatum)等植物组成群丛,表现出极强的耐盐性[13]。研究表明,一定量Na+的参与可促进黑果枸杞的生长[12]。这一发现在本研究中得到了验证,50 mmol/L NaCl处理显著提高了黑果枸杞的鲜重、干重和相对生长速率。在其他旱生植物和盐生植物中也发现类似结果,如霸王(Zygophyllum xanthoxylum)和滨藜(Atriplex patens)[14]。此外,本研究发现,50 mmol/L NaCl处理可促进黑果枸杞种子的萌发,同时明显增加子叶直径及初根长度(P<0.05)。Ma等[15]和Slama等[16]研究发现,在干旱环境中,霸王和马齿苋(Sesuvium portulacastrum)能够吸收大量的Na+并将其作为主要的渗透调节物质。本研究中,50 mmol/L NaCl处理下黑果枸杞中Na+浓度显著增加(P<0.05),进一步表明适当浓度的NaCl添加可通过引起植物中大量Na+的积累,促进植物的生长和发育。K+是引起保卫细胞渗透势变化的重要离子,参与调节细胞吸水和气孔运动的生理过程[17]。在盐碱条件下,维持植物组织中K+的能力是植物耐盐的重要机制[18]。在胁迫条件下,K+含量通常会降低,尤其是当植物中含有大量Na+时,Na+等盐离子直接与K+相互作用或阻碍K+的吸收和运输[19]。本研究表明,与对照相比,添加NaCl虽然降低了叶中的K+浓度,但根与茎中的K+浓度在50和100 mmol/L NaCl处理下明显增加(P<0.05),即使200 mmol/L NaCl处理下有所下降,300和400 mmol/L NaCl处理下又有所提高。推测黑果枸杞植株根系通过吸收大量的K+,一方面满足植株快速生长和发育所需,另一方面以此提高K+/Na+来增加其耐盐性,因此,适当盐胁迫下根部K+吸收率增加[20]。由此,推测NaCl处理可使植株显著积累Na+并保持K+稳定,这可能是50 mmol/L NaCl促进黑果枸杞生长的主要原因。
3.2 NaCl可通过提高光合作用促进植株发育
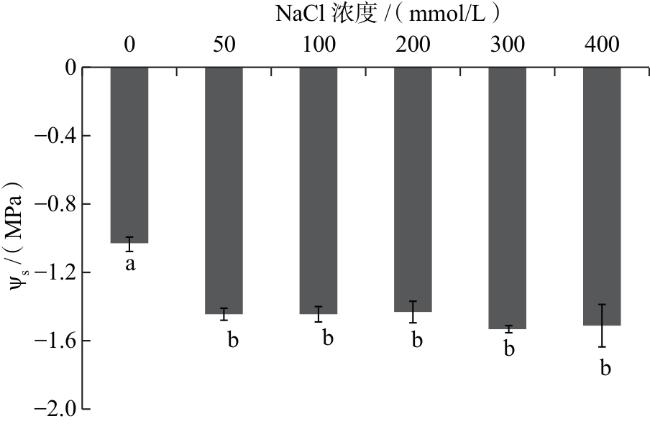
耐渗透胁迫能力强的植株,在盐胁迫下能够维持较高的气孔导度,使CO2同化率处于较高水平[24],从而保障生长叶片获得充足的碳供应[25]。研究表明,叶片的光合速率与气孔导度呈正相关[16,26],而气孔导度依赖于气孔保卫细胞的开放程度[27]。本研究中,NaCl处理下黑果枸杞的净光合速率和相对生长速率与气孔导度均呈极显著正相关,推测NaCl对黑果枸杞生长的刺激作用在一定程度上归因于其促进了气孔保卫细胞的开放。此外,NaCl的加入提高了气孔导度,使黑果枸杞植株的蒸腾能力显著增加,进而影响水分吸收能力,这可能也是盐条件下促进植株生长的原因之一。渗透调节能力作为促进气孔保卫细胞开度的另一因素,是植物抵御逆境环境的重要表现。植物通过主动积累溶质以提高细胞液浓度,进而使渗透势降低,以此维持体内水分平衡,实现对逆境胁迫的适应[28-29]。与对照相比,50 mmol/L NaCl处理的叶中Na+浓度提高,植株的渗透势降低,进而增强了植株的光合作用。
3.3 叶中积累Na+是黑果枸杞抵御盐胁迫的有效策略
盐生植物可分为3种类型[30]:具有泌盐能力的泌盐植物,此类植物一般有盐腺;拒绝盐分吸收和运输的拒盐植物;真盐生植物(也称聚盐植物或稀盐植物),借助从高盐浓度土壤中获取水分,并利用细胞内液泡膜上大量的转运蛋白,将胞质中过量的Na+转至液泡中隔离起来,进而使自身体内的盐浓度维持在较低水平。本研究发现,黑果枸杞各组织中的Na+浓度在正常条件下(CK)无明显差异,当加入50~300 mmol/L NaCl后,叶中的Na+浓度显著高于根和茎,说明盐条件下其具有将Na+积累至叶中的功能。受到盐胁迫时,黑果枸杞可能将Na+积聚到叶细胞的液泡中,以避免离子在细胞质、线粒体、叶绿体等重要细胞器中过量积累,进而维持其正常生理活动和光合作用,这可能是黑果枸杞抵御盐胁迫的有效途径。
综上,本研究表明,适量的NaCl处理可通过提高植株Na+、K+含量及光合能力促进黑果枸杞的生长,并通过将Na+积累至叶中来抵御盐胁迫。