

UV-B辐射是一种影响植物生长发育和环境适应的非生物胁迫因子,在植物的分子遗传、生理生化和表型性状水平上引起一系列复杂的变化。在气候持续变化的背景下,了解植物在紫外光环境下的反应尤为重要。Mckenzie等[1]研究指出,UV-B被视为一种环境信号,是一种影响植物生长的非生物胁迫因子。UV-B辐射能够调节植物光形态建成的各个方面,包括抑制植物下胚轴伸长、子叶扩张以及类黄酮物质的积累等[2-3],从而引发植物的一系列生理应激响应。然而,持续高强度的UV-B辐射可导致植物DNA损伤、氧化应激以及脂质和蛋白质的氧化[4],最终可能导致植物生长发育异常。为了应对这些负面影响,植物采取了多种适应策略,包括形态结构的变化、内源性大分子的合成以及次生代谢物质的产生,以修复DNA损伤和解毒活性氧,减少UV-B辐射诱导的损伤,并增强植物对高强度UV-B辐射环境的适应能力。本文阐述了UV-B辐射对植物形态、生长发育、生理生化过程的影响,UV-B与其他环境因子的复合胁迫,以及植物响应UV-B胁迫的分子机制,同时对UV-B辐射对植物的损伤机理及其应对机制进行展望。
1 UV-B辐射对植物形态及生长发育的影响
1.1 形态发育调控
UV-B辐射能够诱导植物形态发生改变,如叶片卷曲、叶面积减小、叶片厚度增加等。随着UV-B 辐射强度增强,植物子叶下胚轴生长会受到抑制,出现节间缩短,腋生枝条数量增加,根冠比改变等现象[5]。这些形态的调整一定程度上对植物的生长发育起到了保护作用。UV-B辐射可使蚕豆[6](Vicia faba)、黄瓜[7](Cucumis sativus)的茎和叶柄长度缩短以及叶面积明显减小,而大豆(Glycine max)、小麦(Triticum aestivum)等作物出现不同程度的矮化现象。特别是小麦,其节间长随着位置的下降显著缩短,从而显著影响生物量的增加[8]。此外,UV-B辐射对种子萌发也有显著影响,可使种子胚根和下胚轴畸形,抑制根的伸长和芽的生长,最终降低种子萌发率[9]。不同植物响应UV-B辐射的形态发育时期存在差异,如美丽风毛菊(Saussurea superba)和麻花艽(Gentiana straminea)在UV-B辐射过程中株高呈持续降低趋势,而矮蒿草(Kobresia humilis)在UV-B辐射初期和后期株高降低,垂穗披碱草(Elymus nutans)则在辐射初期株高降低[10-11]。UV-B辐射导致的株高降低被视为植物的一种自我保护机制,且植物体内源性生长物质的含量和分布会发生改变,如生长素(IAA)、赤霉素(GA)、乙烯(ETH)。UV-B辐射能够促进植物生长素的分解转化,产生多种氧化产物,这些物质能抑制植物茎的伸长,最终导致植株变矮[12]。UV-B辐射对植物的影响是多方面的,高强度的UV-B辐射会损伤植物DNA结构,影响植物体的膜系统、渗透调节系统、抗氧化防御系统、光合特性和次生代谢物质的产生等,从而对植物的形态发育及生长代谢产生影响,如生理生化异常、物质代谢紊乱,进而使植株的株高、节间长度等发生改变。为适应UV-B辐射胁迫或其他逆境环境的影响,植物体通过改变其形态结构实现自我保护。
1.2 细胞膜系统稳定性
膜系统作为细胞的重要组成部分,承担着隔离胞内与外界环境的功能,并参与物质运输、能量交换和信息传递等过程。高强度的UV-B辐射可导致植物细胞膜系统稳定性降低,甚至破坏膜结构,如膜的组成成分磷脂和不饱和脂肪酸遭到破坏,引发膜脂过氧化现象;叶绿体类囊体的片层结构和叶绿体膜结构也会遭到破坏。此外,植物的生理代谢过程及相关酶结构也可能遭到破坏[13],从而加剧膜系统的损伤。例如,UV-B 辐射可导致紫花苜蓿(Medicago sativa)[14]、垂穗披碱草(Elymus nutans)[15]、杧果(Mangifera indica)[16]等植物体内的丙二醛(Malondialdehyde,MDA)含量升高,抗氧化酶活性下降[17],脂质过氧化程度加深[18],进而导致植物细胞膜的通透性改变,影响植物体细胞与外界环境进行物质运输和信号传导[19];UV-B辐射胁迫下,植物体内会积累大量的MDA,使细胞质膜遭到破坏,导致质膜的通透性增大,这是植物受到UV-B辐射损伤的原因之一。
在植物抗逆性研究中,通过测定植物细胞的膜透性以及植物叶片的相对含水量,来评断植物遭受的胁迫程度。植物叶片的相对含水量反映了植物在蒸腾时对水分亏缺的恢复能力。王红等[20]利用电导率法测定不同UV-B辐射强度下杧果细胞膜的透性和损伤率,发现UV-B辐射增强,电导率和细胞的损伤率增大,叶片的相对含水量降低。邱文静等[15]、刘茜等[11]在垂穗披碱草抗UV-B辐射筛选评价中指出,强抗UV-B辐射植物的相对含水量及细胞膜系统指标(MDA含量、电导率)变化幅度较小。在UV-B辐射胁迫下,相对含水量和电导率之间呈显著负相关,相对含水量下降越慢、电导率上升越慢的植物,能更好地维持质膜结构的完整性和稳定性,从而表现出更强的逆境适应能力。
1.3 渗透调节
植物通过动员渗透调节系统对UV-B辐射胁迫作出应答,当胁迫水平在植物可耐受阈值内,植物细胞会产生多种有机溶质来抵御胁迫。这些有机溶质中,小分子有机溶质如可溶性糖、脯氨酸和可溶性蛋白等物质的积累对渗透调节系统具有显著调节作用,表现为有益于植物的生长发育[21]。研究发现,在逆境胁迫下,植物的渗透调节系统会降低细胞的渗透势,其中可溶性糖的累积在此过程中发挥了重要作用,其有助于维持细胞膨压,从而保障植物能够持续从外部环境吸收水分[22]。黄志慧等[23]、邱文静等[15]研究发现,植物通过积累可溶性糖以响应UV-B辐射和适应逆境环境。陈艳等[24]在不同剂量UV-B辐射与辣椒(Capsicum annuum)渗透调节关系研究中得出,高剂量UV-B辐射显著减少了紫外敏感型辣椒中的可溶性糖含量,而低剂量的UV-B辐射能大幅度提升可溶性糖含量。因此,在植物抗逆性研究领域,可溶性糖含量的累积被视为植物抵抗UV-B辐射的一项重要指标。
植物遭受逆境胁迫时,细胞内的游离脯氨酸合成能力加强,脯氨酸含量的增加被认为是植物受到逆境伤害的重要表现之一[29]。Vives-peris等[30]在柑橘抗UV-B辐射研究中发现,抗UV-B辐射能力强的品种脯氨酸含量积累更快、更高。这与垂穗披碱草[11,15]和罗布麻(Apocynum venetum)[31]响应UV-B辐射的研究结果一致;同时,在UV-B辐射条件下,脯氨酸含量显著上升,其对渗透调节系统的调节作用尤为显著。抗UV-B辐射能力强的植株体内脯氨酸含量比抗UV-B辐射能力弱的植株高1.58倍[30],表明植物抗UV-B辐射的能力与脯氨酸累积量呈正相关;这与谢博勋[31]对罗布麻渗透胁迫下脯氨酸含量以及质膜相对透性的变化研究结果一致。由此表明,植物遭受逆境胁迫时,脯氨酸的生成量与植物抗UV-B能力有关,且其积累量与胁迫持续时间亦存在相关性。
1.4 抗氧化防御
在UV-B胁迫下,植物抗氧化防御机制会激活一系列反应以清除细胞内自由基,因此,自由基清除系统是植物抵御UV-B辐射的关键防御策略。自由基清除系统主要有两类:一类是包括谷胱甘肽(Glutathione,GSH)和抗坏血酸(Ascorbic acid,AsA)等在内的抗氧化剂;另一类是由超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化氢酶(Catalase,CAT)、抗坏血酸过氧化物酶(Ascorbate peroxidase,APX)和过氧化物酶(Peroxidase,POD)、谷胱甘肽过氧化物酶(Glutathione peroxidase,GSH-Px)等组成的抗氧化酶系统。这些物质能够清除细胞内的活性氧自由基,从而有效防止植物膜脂过氧化,以减轻UV-B辐射对植物造成的损伤[32-33]。
植物在遭受逆境胁迫时,其抗氧化防御系统中的抗氧化酶活性会显著提高,从而有效清除植物体内的超氧阴离子,以抵御或修复自由基对细胞造成的损伤。不同强度的UV-B辐射会导致植物体内SOD、POD和CAT活性的差异变化。习亚君[17]研究发现,低剂量的UV-B辐射对叶片中SOD、POD和CAT活性的影响不显著,而高剂量或长时间的UV-B辐射则会严重影响这些酶的活性,且POD活性随着UV-B辐射强度增强逐渐升高,这与任举[34]、赵欣悦等[35]、吕杰等[36]的研究结果一致。Vives-peris等[30]研究表明,抗UV-B辐射能力强的植株和UV-B敏感型植株体内CAT活性无显著差异。然而,大量研究证实,抗逆性强的植物较抗逆性弱的植物,其SOD、POD和CAT等抗氧化酶的活性更强[15,29,34]。
植物遭受UV-B胁迫时,APX和GSH-Px通过催化还原H2O2,以降低过氧化物及其他产物对植物细胞的毒害,使植物抗UV-B辐射能力增强。吕杰等[36]等在不同时长的UV-B辐射烟草(Nicotiana tabacum)、麻花艽等研究中得出,APX活性随着UV-B辐射增强而上升,这与邱文静等[15]对UV-B辐射胁迫垂穗披碱草的研究结果一致。然而,刘英等[37]对UV-B胁迫下杜仲(Eucommia ulmoides)叶片生理特性的研究发现,叶片中APX活性随辐射强度增强呈下降趋势。GSH-Px存在于植物体各个组织中,GSH-Px活性较高时,H2O2分解更快。褚润等[38]研究表明,随着UV-B辐射强度和持续时间的增加,香蒲(Typha orientalis)叶片中GSH-Px活性显著增加,这与Dias等[39]对小麦叶片在UV-B辐射胁迫下的研究结果一致。而邱文静等[15]研究发现,UV-B辐射胁迫下,垂穗披碱草叶片中GSH-Px的活性降低,这与刘茜等[11]的研究结果一致。UV-B辐射能够诱导植物抗氧化剂含量和抗氧化酶活性的提升,从而增强植物的抗氧化能力,以抵御UV-B辐射造成的损伤;然而,当UV-B辐射强度超过植物抗氧化系统的阈值时,其防御能力有限,导致抗氧化物质和抗氧化酶的合成受阻,抗氧化能力下降,最终可能对细胞产生毒害作用。
在UV-B胁迫下,除各种抗氧化酶发挥重要作用外,植物细胞内的非酶类抗氧化保护剂同样发挥着至关重要的作用。AsA和GSH是重要的抗氧化剂和自由基清除剂,能直接清除O3,还可以作为酶反应底物清除活性氧,以减轻氧化胁迫对植物的伤害[38]。随着UV-B辐射剂量增加,植物体AsA含量增加[38],Dias等[39]研究发现,随着UV-B辐射强度增加,小麦叶片细胞中AsA和GSH 含量均有提高。UV-B辐射可导致植物体内活性氧水平增加,同时诱导抗氧化酶活性和抗氧化物质含量相应提升。UV-B辐射会导致植物体内抗氧化酶活性和抗氧化物质含量提升,但在抵御逆境胁迫过程中,尚无法确定具体哪种抗氧化物质发挥主导作用。
1.5 次生代谢
类黄酮化合物是一类广泛存在于植物体内的次生代谢产物,根据其化学结构和性质可划分为酚类、萜类和含氮有机物三类[40]。这些次生代谢产物通过两个方面发挥防御作用:一是作为淬灭剂,能够有效清除活性氧自由基;二是作为过滤剂,可以高效吸收到达叶面的UV-B辐射。UV-B辐射可以诱导类黄酮生物合成途径中相关基因的表达,并促进具有紫外线吸收功能的类黄酮物质在植物体表的积累[41]。这些紫外吸收物质在植物表面积累,有效降低了UV-B辐射的透过率,从而减轻UV-B辐射对叶肉细胞的损伤。UV-B辐射通过诱导调控MYB、bHLH、WD40等基因表达,来调节结构基因的表达[42],进而使植物体内总黄酮含量增加[43-46]。然而,随着胁迫时间延长或胁迫强度增强,光合结构和DNA结构遭到损伤,从而抑制了多种酶的活性表达,导致总黄酮被逐渐分解[47]。此外,花青素作为类黄酮化合物的一种,在植物抵御和适应UV-B辐射胁迫中也发挥着重要作用。UV-B辐射强度增强会诱导植物结构基因和调节基因的表达,进而调控花青素的合成[48]。Zhang等[49]连续4代对水稻进行自然光和UV-B紫外光照射,发现UV-B辐射影响种子形态,使植物体内原花青素含量显著增加,这与Gong等[50]、Liu等[40]在UV-B辐射对植物次生代谢物质含量的影响研究结果一致。类黄酮和花青素等次生代谢物质的积累有助于植物免受或减轻UV-B辐射引起的损伤。
1.6 光合作用
叶片作为植物进行光合作用的主要场所,其叶绿素含量可作为衡量UV-B胁迫对植物造成损伤的一个重要指标。UV-B辐射胁迫下,植物体会通过提升叶绿素含量来抵御胁迫;然而,UV-B辐射强度过高时,植物的自身防御机制被激活,导致PS II最大光化学效率降低,进而影响光合速率。在不同的UV-B辐射强度下,植物叶片光合色素含量以及光合特性存在差异[51]。UV-B辐射会导致美丽风毛菊叶片中叶绿素和类胡萝卜素含量降低,进而影响叶片光能吸收和传递,并降低原初光化学反应的效率[52],这可能是UV-B辐射阻碍了植物叶绿素的合成或叶绿素合成后被降解。UV-B辐射胁迫下,老芒麦(Elymus sibiricus)的光合速率变化极为敏感[53]。王林叶等[54]在野生型籼稻两用不育系812 S和光氧化突变株系812 HS的研究中得出,UV-B辐射增强,光氧化突变株系812 HS的PS Ⅱ最大光化学效率在分蘖期的提高幅度比野生型812 S水稻更大,而抽穗期和灌浆期则明显低于野生型812 S;PS Ⅱ最大光化学效率失活的原因是质体醌PQs和PSI蛋白复合体对UV-B辐射敏感,电子从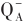 向
向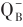 传递过程受阻,致使快还原型库PQ受影响。由此表明,适量UV-B辐射有益于植物叶片叶绿素的合成,从而增强光合作用;而UV-B辐射持续时间或强度超过阈值时,植物PS Ⅱ最大光化学效率受损,影响叶绿素和光合蛋白复合体的合成,进而抑制植物的光合作用。
传递过程受阻,致使快还原型库PQ受影响。由此表明,适量UV-B辐射有益于植物叶片叶绿素的合成,从而增强光合作用;而UV-B辐射持续时间或强度超过阈值时,植物PS Ⅱ最大光化学效率受损,影响叶绿素和光合蛋白复合体的合成,进而抑制植物的光合作用。
向 传递过程受阻,致使快还原型库PQ受影响。由此表明,适量UV-B辐射有益于植物叶片叶绿素的合成,从而增强光合作用;而UV-B辐射持续时间或强度超过阈值时,植物PS Ⅱ最大光化学效率受损,影响叶绿素和光合蛋白复合体的合成,进而抑制植物的光合作用。2 植物响应UV-B胁迫的分子机制
2.1 DNA损伤修复与基因表达调控
UV-B辐射通过作用于碱基和多核苷酸链影响植物DNA的结构[55]。植物自身具备碱基切除修复和光复活修复等机制,但其修复能力受限于胁迫强度。当辐射剂量超过阈值时,修复效率显著下降,最终导致细胞程序性死亡[55]。多组学联合分析(如转录组与代谢组)揭示了UV-B胁迫下基因表达的动态重编程特征。高海拔植物中光反应关键基因rbcL的表达随UV-B强度增加呈现梯度抑制,极端胁迫下其表达几乎完全关闭,导致光合效率下降和形态发育异常[56]。同时,抗性相关基因的激活可显著提升植物的耐受性,如水稻OsRLCK160受体激酶与OsbZIP48转录因子通过磷酸化级联调控类黄酮合成通路,并通过磷酸化正向调节植株中类黄酮的积累,从而提高水稻对UV-B辐射的抗性[57];矮牵牛(Petunia hybrida)中F3H基因的高表达则通过增加槲皮素含量增强光保护能力[58]。这些研究表明,基因表达调控网络在权衡DNA修复与代谢资源分配中起核心作用。
2.2 UVR8信号通路及其互作网络
紫外光受体UVR8是植物感知UV-B信号的核心元件。其以同源二聚体形式存在,UV-B诱导色氨酸残基(W233/W285/W337)光解后单体化,进而与COP1-SPA复合物互作,抑制E3泛素连接酶活性以稳定转录因子HY5,促进光形态建成及次生代谢物积累[59]。负反馈调节因子RUP通过竞争性结合UVR8单体,加速其二聚化以终止信号传递,形成动态平衡[60]。结构生物学研究进一步揭示,COP1的WD40结构域通过双界面结合UVR8单体,其中R420残基对复合物稳定性至关重要;而RUP2与UVR8的互作则依赖D129残基,并通过增强氢键网络促进UVR8从COP1解离[59,61]。此外,UVR8介导的信号转导途径在植物响应UV-B胁迫中起到多种作用,如抑制叶肉细胞伸展、下胚轴伸长和提高光合速率等功能及对各种植物激素的响应[62-64]。该通路不仅调控下胚轴伸长抑制和光合适应,还通过互作网络影响激素信号(如茉莉酸)的合成,形成多维防御网络[59]。
2.3 激素调控与次生代谢基因激活
植物通过激素信号与次生代谢协同响应UV-B胁迫,油菜素甾醇(Brassinolide,BR)信号在此过程中发挥枢纽功能。BR激活的转录因子BES1通过与BIM1互作,抑制黄酮醇合成基因MYB11/12/111的表达,优先维持植物生长相关代谢[65-66]。UV-B辐射通过两种途径重塑该调控网络,一方面,UVR8单体直接结合BES1,阻断其与靶基因的结合,解除对MYB基因的抑制,促进类黄酮和花青素等光保护物质的合成;另一方面,UV-B还可绕过UVR8直接下调BES1活性,迫使植物从生长模式切换至防御模式[65-67]。进一步研究表明,UV-B激活的UVR8-TCP4复合物直接结合茉莉酸(Jasmonic Acid,JA)合成基因LOX2的启动子,显著上调JA含量,进而增强花青素通路活性并降低ROS毒性[59]。这一机制在青藏高原抗UV-B蔓菁品种KTRG-B48b中得到验证,其通过快速诱导JA合成,有效清除过氧化氢并提升耐受性[59,61]。激素与次生代谢的联合调控,揭示了植物通过分子开关(如BES1)和代谢重编程(如JA和花青素等)实现多层次适应性响应的策略[67]。
3 UV-B辐射复合胁迫与生态适应
植物所处的生存环境具有复杂性,其所受到的胁迫也具有多样性,UV-B辐射与其他环境因子的复合胁迫研究逐渐成为热点。目前,关于UV-B辐射与复合胁迫的研究主要集中在两个领域:一是与环境因素和营养因素的交互作用,如温度、干旱、土壤盐碱程度、氮和外源激素等;二是与污染因子的联合效应,如重金属、酸雨以及病原微生物等。韩超等[68]在UV-B辐射胁迫下研究云杉(Picea asperata)幼苗的光合及生理响应发现,升高温度能缓解UV-B辐射胁迫对云杉光合作用的损伤,植物的生理生化特性也发生了明显变化。郭晓瑞等[69]对氮素与UV-B辐射复合作用的研究表明,氮含量的增加能够为长春花(Catharanthus roseus)合成用于抵抗UV-B辐射的次级代谢产物提供原料,进而缓解UV-B辐射对其生长的抑制作用。这与张来军等[70]研究植物对褪黑素与UV-B复合胁迫的响应结果一致,即外源褪黑素对UV-B辐射造成的DNA损伤有一定的修复作用。此外,部分因子与UV-B辐射复合胁迫具有增强协同损伤的效应。如UV-B辐射与酸雨复合胁迫对大豆的生长具有协同抑制作用[71]。高潇潇等[72]研究两种水稻品种幼苗对UV-B辐射和接种稻瘟病菌(Magnaporthe grisea)的生理响应,表明胁迫因子的作用顺序、稻瘟病株、UV-B辐射剂量对不同水稻品种的表现和生理指标变化趋势均存在差异。因此,UV-B辐射胁迫或与其他因子的复合处理对植物的影响通常表现为协同或拮抗效应,与所考察指标的性质密切相关。
4 结语
UV-B辐射对植物的生长发育、形态结构、生理生化过程及分子遗传产生了广泛影响,而这些复杂变化往往与植物的关键功能紧密相关。高强度UV-B辐射会损伤植物的物质与能量代谢,干扰植物体内信号转导与信息传递机制,从而对植物生长发育及形态建成产生影响。相反地,植物体能够通过改变自身形态结构来实现自我保护。
近年来,关于UV-B辐射对植物造成的损伤和植物自我防御机制是研究热点之一。由于植物响应UV-B辐射的复杂性,目前研究主要集中于农作物和草本植物,林木类植物的相关研究较少;另外,大多数研究在人工气候箱和温室内进行,难以真正模拟自然环境,还需通过野外自然环境进一步验证;植物对UV-B辐射的响应和适应是长期积累的过程,瞬时或短期的结果不能完全诠释植物对UV-B辐射的长期适应。因此,植物响应UV-B辐射机制的研究有待于进一步加强,尚需从分子遗传方面阐明UV-B辐射对植物生理生化过程的影响。在分子生物学飞速发展的当下,UV-B辐射对植物造成损伤的分子机理将会更明晰和具体,这将有助于有针对性和高效地应对UV-B辐射对植物的负面影响。