

金顶侧耳(Pleurotus citrinopileatus)是一种珍贵的食、药用真菌,主要分布于云南、广西、四川等地[1]。其凭借高蛋白、低糖、低脂的营养特性,在膳食结构中占据重要地位。Zhang等[2]研究表明,金顶侧耳具有较高的药理活性,其子实体多糖具有免疫增强作用。王沛等[3]通过液体摇瓶培养技术提取的菌液,有效促进了小鼠体液免疫与细胞免疫功能的提升。随着食用菌全基因组序列的解析,利用现代分子生物学手段调控其多糖合成途径中的关键基因,已成为提升多糖产量的重要策略之一[4-6]。
磷酸葡萄糖变位酶(Phosphoglucomutase,PGM)的核心功能是催化葡萄糖-6-磷酸与葡萄糖-1-磷酸之间的可逆转化。其中,葡萄糖-6-磷酸进入分解代谢过程以产生能量,而葡萄糖-1-磷酸是糖核苷酸的前体,被细胞用于合成各种多糖。PGM在细胞壁和多糖生物合成中具有重要功能[7]。例如,在灵芝中过表达α-PGM,可显著增强PGM及磷酸葡萄糖异构酶基因的转录水平及相关酶活性,从而提高灵芝多糖含量[4]。目前,研究人员已在拟南芥(Arabidopsis thaliana)[8]、玉米(Zea mays)[9]、马铃薯(Solanum tuberosum)[10]、广东虫草(Cordyceps guangdongensis)[11]等植物和真菌中克隆出PGM基因。然而,有关金顶侧耳PGM基因的相关研究较少。为此,本研究以金顶侧耳全基因组数据为基础,通过特异性引物对PGM基因进行克隆与生物信息学分析,为其在食、药用方面的应用提供参考。
1 材料与方法
1.1 试验材料
金顶侧耳由蚌埠学院食品与生物工程学院提供。取健康菌丝放于离心管中,液氮速冻并于-80 ℃冰箱保存,用于总RNA提取。
1.2 试验方法
1.2.1 总RNA的提取和反转录
收集金顶侧耳新鲜菌丝,取50~90 mg放入灭菌研钵中,加液氮研磨成粉末状,按照总RNA快速提取试剂盒(Easy Plant RNA Extraction Kit)说明书提取金顶侧耳总RNA,并按照cDNA逆转录试剂盒说明书合成cDNA。逆转录反应体系为20 μL,即1 μL金顶侧耳总RNA;4 μL 5×Polestar RT Master Mix (with dsDNase);14 μL Rnase Free H2O;1 μL 20×Oligo dT (25) & Random Primer。合成的cDNA稀释3倍于-20 ℃冰箱保存,用于后续试验。
1.2.2 PGM的克隆与鉴定
根据金顶侧耳基因组中已获得的PGM基因序列,设计引物PGM-F:ATGGATTCCTTGCGCCCATTGG;PGM-R:TCAAGGCTTTCCAAGGGAGTGT,并送至生工生物工程(上海)股份有限公司合成。PCR扩增体系为25 μL(表1)。PCR反应体系:95 ℃预变性3 min;95 ℃变性15 s;54.5 ℃退火15 s;72 ℃延伸30 s,35个循环。反应结束后,PCR产物进行1%琼脂糖凝胶电泳。目的基因胶回收采用北京百泰克生物科技有限公司柱式Column DNA Back试剂盒,后连接pMD 19-T载体并转化Escherichia coli DH5α。挑取阳性克隆凝胶电泳后,测序鉴定目的片段。
表1 PCR反应体系 (μL) |
| 组分 | 体积 |
|---|---|
| ddH2O | 9.5 |
| PGM-F | 1 |
| PGM-R | 1 |
| cDNA | 1 |
| 2×Rapid Taq Master Mix | 12.5 |
1.2.3 生物信息学分析
采用DNAMAN软件进行基因片段拼接与氨基酸同源性多重比对;Expasy-ProtParam在线工具分析PcPGM蛋白的理化特性;Prot Scale在线程序分析亲疏水性;TMHMM软件分析蛋白跨膜域;SignalP软件进行信号肽预测;NetPhos 3.1软件预测磷酸化位点;ProtComp 9.0软件预测亚细胞定位;SOPMA软件进行PGM基因编码氨基酸序列的二级结构预测;SWISS MODEL软件预测蛋白三级结构;MEGA软件构建系统进化树;String软件预测蛋白互作网络。
2 结果与分析
2.1 PGM的克隆与鉴定
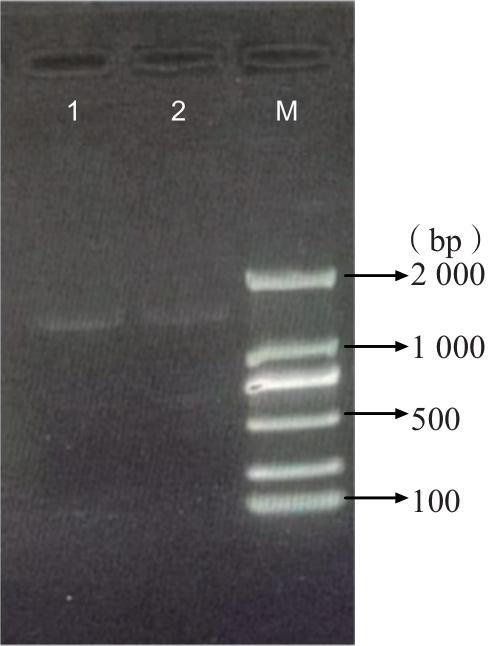
以金顶侧耳逆转录后稀释的cDNA为模板,采用PGM-F/R引物对其进行PCR扩增。1%琼脂糖凝胶电泳结果显示,在1 800 bp处出现单一的目标条带(图1)。进一步的测序分析得到的开放阅读框长度为1 797 bp。表明已成功克隆了PGM基因的cDNA全长,并命名为PcPGM。
2.2 理化性质
由表2可知,PcPGM基因编码的蛋白质由598个氨基酸组成,分子量为66.35 kDa,等电点为5.98,是一种酸性蛋白质。其中带正电荷的氨基酸残基(天冬氨酸+谷氨酸,Asp+Glu)为73,带负电荷的氨基酸残基(精氨酸+赖氨酸,Arg+Lys)为63。亲水性(GRAVY值)平均值为-0.278,预测该蛋白为亲水性蛋白。该蛋白的不稳定指数为29.54,预测该蛋白为稳定蛋白。
表2 PcPGM蛋白的理化性质 |
| 预测值 | PcPGM |
|---|---|
| 氨基酸数量 | 598 |
| 蛋白分子量/kDa | 66.35 |
| 理论等电点 | 5.98 |
| 分子式 | C2978H4623N793O884S21 |
| GRAVY值 | -0.278 |
| 不稳定指数 | 29.54 |
| Asp+Glu | 73 |
| Arg+Lys | 63 |
2.3 亲水性、信号肽和跨膜结构
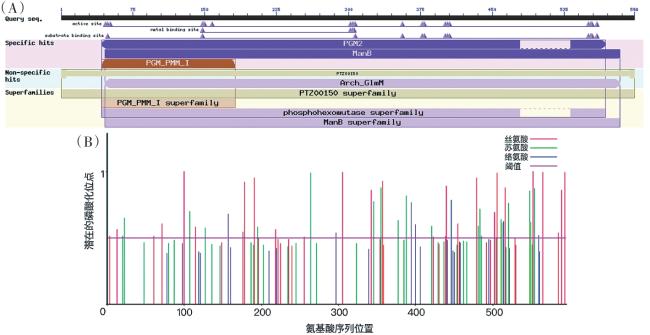
2.4 功能结构域、亚细胞定位和磷酸化位点
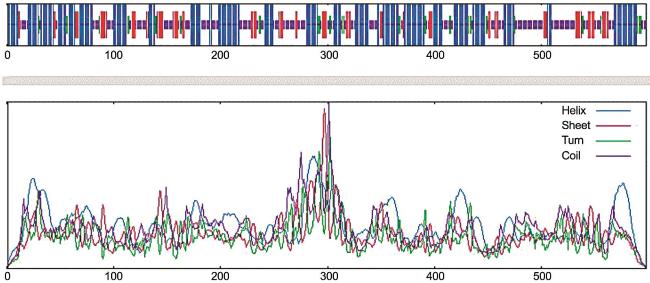
2.5 PcPGM蛋白的二级结构
由图4可知,PcPGM蛋白二级结构中α-螺旋包含258个氨基酸,占总氨基酸的43.14%,无规则卷曲包含214个氨基酸,占总氨基酸的35.79%,延伸链包含88个氨基酸,占总氨基酸的14.72%,β-折叠包含38个氨基酸,占总氨基酸的6.35%。
2.6 三级结构
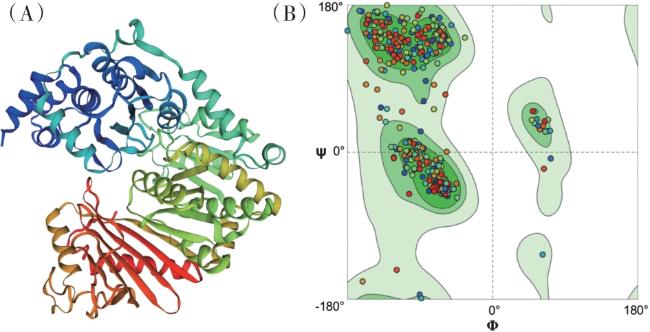
利用SWISS-MODEL在线软件对PcPGM基因编码蛋白的三级结构进行预测分析,共构建出4个PcPCM蛋白三级结构模型图,由图5A可知,其序列与虫拟蜡菌(Ceriporiopsis subvermispora strain B)M2QNP8_CERS8(PGM)蛋白的结构具有较高的相似性,序列一致性达70.39%,表明PcPGM的三级结构模拟的结果具有较高的可靠性。进一步分析显示,PcPGM蛋白的三级结构主要是由α-螺旋和无规则卷曲组成,同蛋白质二级结构预测结果一致。三级结构模型评估结果显示,95.3%以上的氨基酸残基位于可信区域内(图5B),Ramachandran异常值为1.01%,证明了其空间结构较稳定。
2.7 PcPGM蛋白的序列同源性与系统进化树
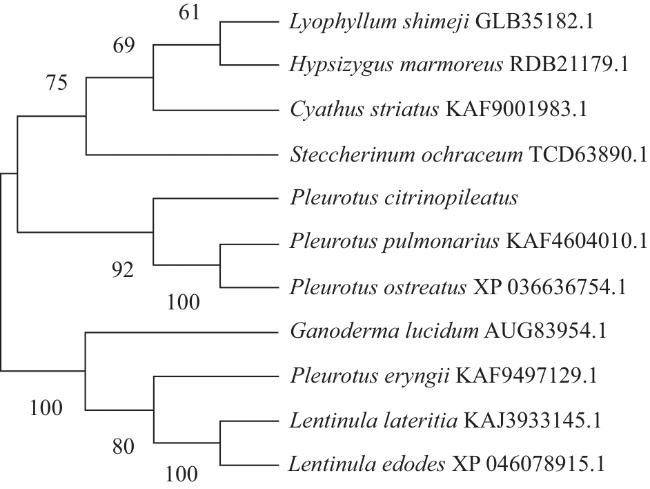
表3 PcPGM蛋白同源序列比对单位:% |
| 物种名 | 相似性 |
|---|---|
| 平菇(Pleurotus ostreatus) | 78.04 |
| 秀珍菇(Pleurotus pulmonarius) | 79.70 |
| 真姬菇 (Hypsizygus marmoreus) | 71.33 |
| 玉蕈离褶伞(Lyophyllum shimeji) | 67.50 |
2.8 蛋白互作网络
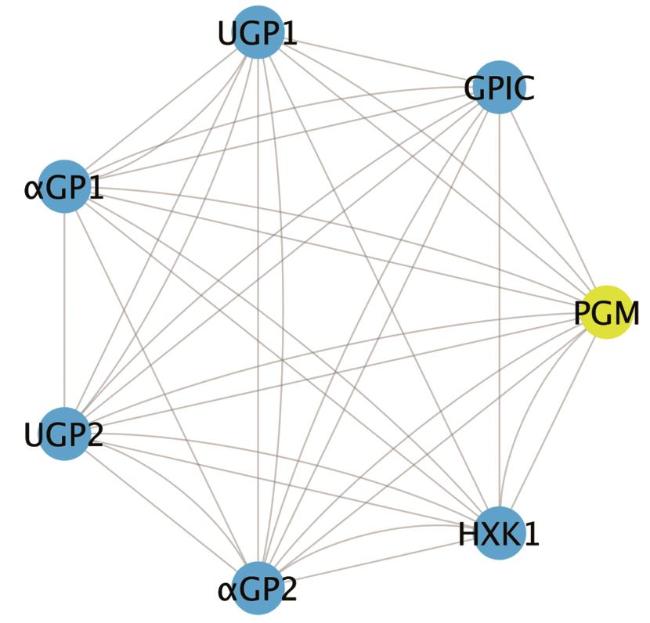
由图7可知,String蛋白互作网络分析共获得7个节点和36条互作关系,与PcPGM蛋白可能存在互作关系的蛋白质共有6个,包括己糖激酶1(Hexokinase-1,HXK1),葡萄糖-6-磷酸异构酶(Glucose-6-phosphate isomerase,GPIC),α-葡聚糖磷酸化酶1(Alpha-glucan phosphorylase 1,αGP1),αGP2,utp-葡萄糖-1-磷酸尿苷酰转移酶1(UTP-glucose-1-phosphate uridylyltransferase 1,UGP1)和UGP2这些蛋白均是糖代谢相关的重要酶类,说明PcPGM在金顶侧耳糖代谢中具有重要作用。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
多糖是自然界中广泛存在的物质,是维持生命活动不可或缺的成分,主要为生物体提供碳源和能量。金顶侧耳是一种具有重要经济价值的食用菌,因其较高的营养成分和药用价值而备受关注。宋泽文[12]研究表明,金顶侧耳子实体、菌丝体或发酵液中均含有丰富的多糖,且其能够治疗尾部悬吊大鼠所致的肌肉萎缩。PGM是糖代谢途径中的一个关键酶,其活性对碳水化合物代谢、能量平衡以及细胞的整体功能具有重要影响[7]。相关研究表明,不同真菌PGM基因序列存在一定的差异性,如茯苓PGM与绣球菌PGM、灵芝PGM、血红栓菌PGM、冬生多孔菌PGM的同源性分别为87%、85%、85%、84%[13]。本研究中的同源序列分析表明,不同真菌间PGM基因序列的保守性相对较高;系统进化树分析可知,不同真菌PGM蛋白的功能在进化上是较为保守的,金顶侧耳与秀珍菇、平菇在进化上处于同一分支上,与金顶侧耳亲缘关系相对较远同属香菇属的香菇和砖红小香菇也处于同一分支,这些均与分类学结果一致。
Li等[14]研究表明,细胞内大多数蛋白质以多聚体形式存在,并在细胞过程中发挥关键的生物学功能。蛋白互作网络分析是从网络层面研究蛋白质功能及其相互作用的有效方法[15]。本研究利用String在线数据库对蛋白质相互作用网络进行分析,结果显示,有6种蛋白质与PGM互作。其中,己糖激酶不仅在糖代谢中起重要作用,还参与调节细胞能量平衡和感应细胞内外糖浓度。此外,己糖激酶1作为一种糖感受器,调节与糖代谢相关的信号传导路径[16]。葡萄糖-6-磷酸异构酶是糖酵解和糖异生途径中的关键酶,催化葡萄糖-6-磷酸和果糖-6-磷酸的相互转化;该酶在能量代谢中发挥重要作用,帮助细胞有效利用葡萄糖[17]。α-葡聚糖磷酸化酶广泛存在于植物、动物和微生物中,其通过催化α-1,4-糖苷键的磷酸解反应,可将葡聚糖链中的葡萄糖单元逐步释放为葡萄糖-1-磷酸[18]。UTP-葡萄糖-1-磷酸尿苷酰转移酶可将葡萄糖-1-磷酸转化为UDP-葡萄糖,在细胞代谢途径中作为葡萄糖基供体发挥核心作用[19]。这些蛋白均是糖代谢的重要酶类,表明PcPGM在金顶侧耳糖代谢中扮演着重要角色。
综上,本研究成功利用金顶侧耳克隆出PGM基因,并运用生物信息学对其进行多角度分析。结果表明,金顶侧耳PGM基因序列全长为1 797 bp,编码598个氨基酸,为稳定、亲水性、非分泌蛋白、定位于细胞质;属于PTZ00150基因家族,具有PGM保守结构域,空间结构较为稳定,进化较为保守;在糖代谢中具有重要作用。本研究为金顶侧耳的糖代谢机制及其育种与应用提供参考。