

褐变是新鲜水果和蔬菜等在加工或贮藏中受到机械损伤后,物质氧化聚合成褐色色素或黑色素的过程,影响商品的营养价值和品质。按发生机理可将褐变分为非酶促褐变和酶促褐变两大类[1]。非酶促褐变是指食品加工和贮藏过程中含氮化合物如胺、蛋白质或氨基酸等与还原糖羰基发生复杂的化学反应,生成褐色物质的现象。褐变程度与丙二醛、脯氨酸和可溶性糖含量密切相关。邢枫等[2]研究认为,采收后的组织衰老、冷害、机械损伤和病原微生物污染等逆境胁迫因素也可加速褐变。Adam等[3]研究表明,褐变是酶催化酚类物质形成醌及其聚合物的结果,与酶活性和多酚类物质的氧化密切相关;酶促褐变多发生在牛油果等果蔬[4-5]和食用菌中。催化酶促褐变的酶有酚氧化酶[6]、过氧化物酶等。酚氧化酶是一类含铜离子的末端氧化酶,包括酪氨酸酶、多酚氧化酶和漆酶等,其在褐变过程中发挥着重要作用。
双孢蘑菇是重要的栽培食用菌之一,其子实体富含蛋白质、多糖和维生素等营养成分[7]。Zhang等[8]研究表明,双孢蘑菇的保质期3~4 d,子实体在采后贮藏及加工过程中极易褐变,是影响鲜销的主要原因之一。金晓燕等[9]对香菇子实体褐变机理进行研究,发现采后的子实体随着贮藏时间的延长,其活跃的后熟过程会引发多种生理生化变化,一些小分子物质和酶的活性发生变化,使菇体组织中的氧化还原平衡遭到破坏。因此,深入了解双孢蘑菇采收后常温储存过程中生理生化变化对认识其褐变机理具有重要意义。
本试验通过测定双孢蘑菇子实体在采收后常温储存下丙二醛、蛋白质、脯氨酸和可溶性糖等的含量,以及酪氨酸酶、多酚氧化酶、漆酶和过氧化物酶的活性,探讨其与褐变的关系,为蘑菇类储藏保鲜以及抑制加工过程中的褐变提供参考。
1 材料与方法
1.1 材料处理
新鲜双孢蘑菇由安徽科技学院食用菌研究所提供。挑选菇体完整、大小均匀、颜色洁白、未开伞、无病虫害且无机械损伤的子实体,洗净晾干后装入塑料筐,用PE保鲜袋套筐但不封口,于常温下储存。每隔1 d随机挑取适量子实体,测量贮藏期间菇体的失重率、褐变度、营养成分含量和酶活性。
1.2 测定指标和方法
1.2.1 失重率计算
测定双孢蘑菇储存前后鲜重,失重率计算如式(1) 。
失重率(%)=(储存前鲜重-储存后鲜重)/储存前鲜重×100
1.2.2 褐变度测定
取1 g双孢蘑菇,加4 mL 0.2 mol/L磷酸缓冲液(pH 6.8,含0.15 mol/L氯化钠,2.5%聚乙烯吡咯烷酮),冰浴研磨,4 ℃离心5 min,转速15 000 r/min,取上清液,于450 nm波长处测定吸光度值,该值用以表示褐变度。
1.2.3 可溶性糖含量测定
取1 g双孢蘑菇,加10 mL蒸馏水,塑料薄膜封口,于沸水中提取30 min,共2次,提取液过滤合并后定容。采用硫酸蒽酮法[10]测定样品在630 nm波长处的吸光度值,根据标准曲线拟合的公式计算可溶性糖含量。
1.2.4 蛋白质含量测定
取1 g双孢蘑菇,加入0.02 mol/L磷酸缓冲液(pH 7.2)10 mL,充分研磨,静置片刻后吸取上清液,8 000 r/min离心10 min。采用Bradford法[11]测定595 nm波长处的吸光度值,根据标准曲线拟合的公式计算蛋白质含量。
1.2.5 脯氨酸含量测定
配制系列浓度脯氨酸溶液,分别吸取2 mL系列浓度的脯氨酸溶液、2 mL冰醋酸和2 mL酸性茚三酮溶液混合,沸水浴中加热30 min,冷却后测定520 nm处吸光度值,制作标准曲线。称取1 g双孢蘑菇,加入3%磺基水杨酸溶液10 mL研磨,在沸水浴中提取10 min,冷却后过滤,滤液即为脯氨酸提取液。吸取适量提取液按照标准曲线方法测得吸光度值,计算脯氨酸含量。
1.2.6 丙二醛含量测定
取1 g双孢蘑菇,剪碎后加入10 mL 10%三氯乙酸和少量石英砂,研磨成匀浆,5 000 r/min离心10 min。吸取上清液2 mL(对照为2 mL蒸馏水),加入0.6 %的硫代巴比妥酸溶液2 mL,沸水浴反应15 min,迅速冷却后离心。取上清液并测定532、600和450 nm波长处的吸光度值,计算如式(2) 。
丙二醛含量(μmol/g)=(A 532-A 600)×6.45-0.56×A 450
1.2.7 多酚氧化酶活性测定
取4 g双孢蘑菇,加入10 mL 0.05 mol/L磷酸缓冲液(pH 6.8,含2.5 %聚乙烯吡咯烷酮),冰浴研磨,4 ℃下6 000 r/min离心15 min,上清液即为多酚氧化酶粗提取液。0.05 mol/L磷酸缓冲液(pH 6.8)1.95 mL和0.1 mol/L邻苯二酚0.5 mL混合后预热至25 ℃,然后迅速加入酶粗提液50 μL混匀,记录420 nm波长处吸光度值在3 min内的变化[12]。
1.2.8 过氧化物酶活性测定
取4 g双孢蘑菇,剪碎后加入10 mL预冷的0.1 mol/L乙酸-乙酸钠缓冲液(pH 5.5,含4%聚乙烯吡咯烷酮)和少许石英砂,冰浴研磨,4 ℃下6 000 r/min离心30 min,上清液为过氧化物酶粗提液。25 mmol/L愈创木酚3 mL、0.5 mol/L H2O2溶液200 μL和0.5 mL酶提取液混匀后立即计时,在反应15 s时记录470 nm处吸光度值作为初始值,每隔30 s记录1次,连续记录6 min,以灭活的酶液作为对照[13]。
1.2.9 酪氨酸酶活性测定
取4 g双孢蘑菇,加入10 mL 预冷的0.05 mol/L磷酸缓冲液(pH 6.8),冰浴研磨成匀浆,4 ℃下6 000 r/min离心20 min,即为酪氨酸酶粗提液。1 mmol/L L-酪氨酸磷酸盐缓冲液(pH 6.8)3.9 mL与0.1 mL酶液迅速混匀,在317 nm处测定吸光度值,连续测定10 min。酶活力单位(U)定义为每分钟引起吸光度值变化0.1所需的酶量。
1.2.10 漆酶活性测定
取4 g双孢蘑菇,加入10 mL预冷的0.05 mol/L磷酸缓冲液(pH 6.8),冰浴研磨成匀浆,4 ℃下6 000 r/min离心15 min,上清液即为漆酶粗提液。0.5 mL 3.36 mmol/L邻联甲苯胺、3.4 mL 0.1 mol/L醋酸缓冲液(pH 4.6)和0.1 mL酶液混匀,25 ℃反应10 min,测定600 nm波长处的吸光度值。
2 结果与分析
2.1 常温储存下双孢蘑菇失重率变化
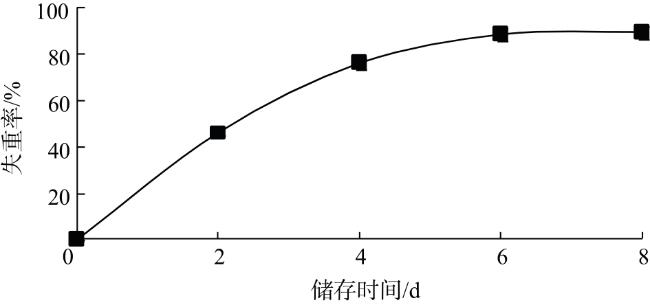
水分含量是衡量双孢蘑菇新鲜程度的重要指标之一。由图1可知,储存6 d时,失重率达88.7%,说明双孢蘑菇采后极易失水。由于双孢蘑菇只有很薄的外皮保护,常温(25 ℃)储存时,失重率呈持续上升趋势,这可能导致其褐变速度加快。
2.2 常温储存下双孢蘑菇褐变度变化
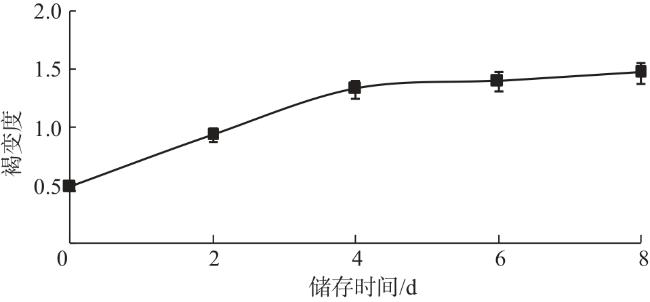
褐变是双孢蘑菇采后品质和商品价值降低的主要原因之一,不仅影响产品外观,而且损害其风味和营养品质。如图2所示,常温储存的前4 d褐变度增加较快,第4天达到1.326,之后褐变速度变慢。在整个储存过程中,双孢蘑菇褐变度持续增加。
2.3 常温储存下双孢蘑菇营养成分含量变化
2.3.1 可溶性糖含量变化
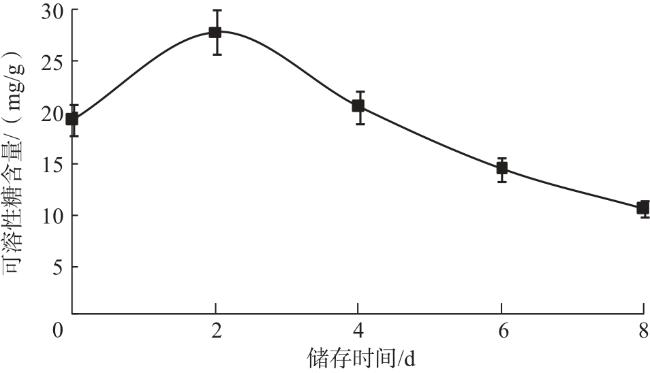
可溶性糖是生物体的重要营养物质之一。试验测定的可溶性糖的标准曲线方程为y=0.014 3x-0.060 0,R 2为0.997 1,由此计算得到不同储存时间的子实体的可溶性糖含量。由图3可知,双孢蘑菇常温储存期间,可溶性糖含量在前2 d呈上升趋势,随后持续下降。
2.3.2 蛋白质含量变化
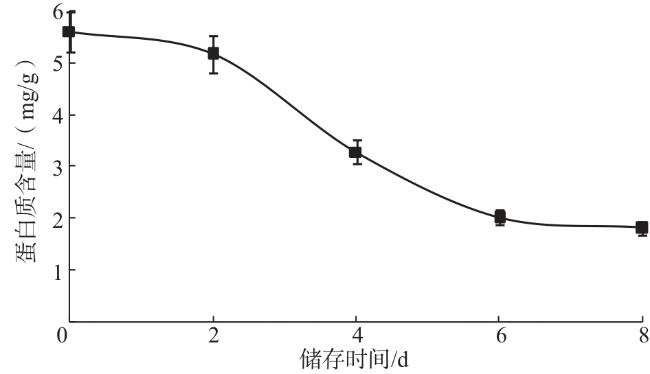
蛋白质是双孢蘑菇子实体的重要组成成分之一。试验测定的蛋白质标准曲线方程为y=0.021 7x-0.017 8,R 2为0.991 3,由此计算得到不同储存时间的子实体蛋白质含量。由图4可知,在双孢蘑菇常温储存1~6 d,蛋白质含量降低速度较快,说明蛋白质的分解代谢可能较为旺盛;常温储存6~8 d,蛋白质含量变化较小。
2.3.3 脯氨酸含量变化
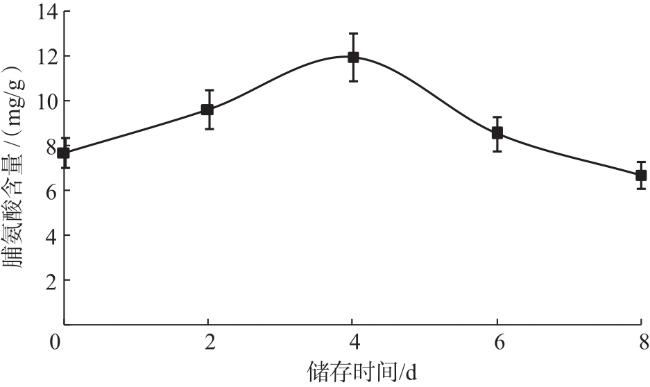
脯氨酸是一种小分子的渗透调节物质,植物在受到逆境胁迫时可能积累高水平的脯氨酸。试验测定的脯氨酸标准曲线为y=0.038 3x-0.013 7,R 2为0.994 6,由此计算得到不同储存时间的子实体的脯氨酸含量。由图5可知,在常温储存的前4 d,双孢蘑菇脯氨酸含量持续上升,随后开始下降。
2.3.4 丙二醛含量变化
2.4 常温储存下双孢蘑菇酶活性变化
2.4.1 多酚氧化酶和过氧化物酶活性变化
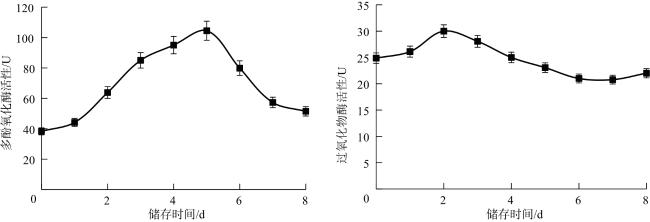
由图7可知,常温储存前期,子实体的多酚氧化酶活性逐渐升高,第5天达到104.509,随后酶活性开始下降,这可能与褐变组织中底物浓度降低或者褐变产物的积累有关。从整体看,多酚氧化酶的活性相对较高,最后降至初始水平,说明其可能在双孢蘑菇褐变过程中发挥主要作用。常温储存2 d内,过氧化物酶活性增强,可能是逆境胁迫刺激细胞产生过氧化物酶来抵御活性氧的伤害。随着储存时间延长,褐变逐渐发生,过氧化物酶活性稍降。说明过氧化物酶持续发挥作用,可能参与双孢蘑菇渐进的褐变过程。
2.4.2 酪氨酸酶和漆酶活性变化
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
双孢蘑菇采收后在常温储存过程中,由于呼吸作用的不断增强,其内含物如水、糖和蛋白质等含量会发生较大变化。其失水后不仅影响正常的生理过程,还会造成失鲜、失重和品质下降。石钰琢等[14]研究发现,失水后双孢蘑菇子实体内相关氧化酶活性增加,呼吸强度增大,从而加快其褐变速度。因此,降低双孢蘑菇的采后失水可以提高其保鲜效果,这与张国强等[15]的研究结果一致。可溶性糖是生物体中的重要碳源,也是响应外界胁迫的诱导分子。王霆[16]研究认为,双孢蘑菇在采收后受干燥空气的胁迫,为减少水分散失而主动积累可溶性糖进行渗透调节,随着时间延长,双孢蘑菇因缺少外界能量的适时补充,呼吸作用持续消耗储存的可溶性糖为其他代谢提供能量,后期可溶性糖含量随贮藏时间延长而降低,本试验结果与此一致。在常温储存过程中,双孢蘑菇蛋白质含量呈下降趋势,说明采摘后其可能受到水分等因素的胁迫而趋于衰老,蛋白质不断被降解。
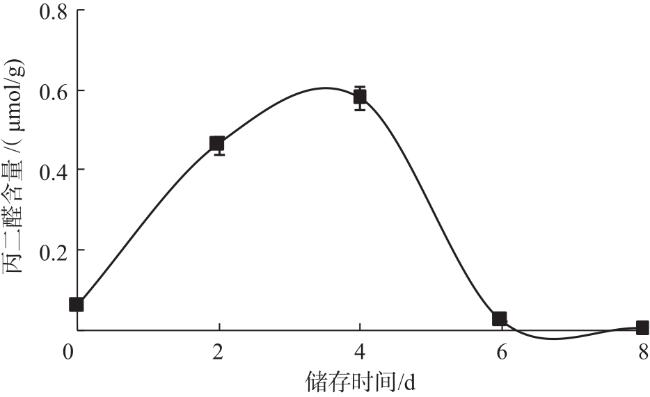
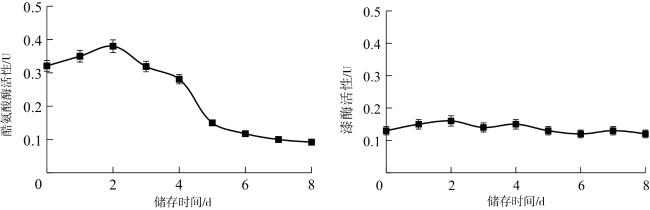
脯氨酸含量呈先上升后下降的趋势,与Acosta-Motos等[17]、Liu等[18]的研究结果一致。在储存初期,双孢蘑菇通过积累大量游离脯氨酸来增强组织的渗透调节能力,维持体内大分子物质的稳定性;随着储存时间的延长,由于缺少外界的能量和物质补充,脯氨酸或被降解用于供能或被用于合成其他蛋白质,其含量逐渐降低。丙二醛含量呈先上升后下降的趋势,说明可能在双孢蘑菇褐变过程中,代谢平衡被打破,活性氧等自由基逐渐累积,细胞受到氧化胁迫,丙二醛含量升高,而后相关抗氧化酶被激活发挥保护作用,从而含量下降[19]。徐耀宗等[20]对低压静电场处理对双孢蘑菇采后贮藏品质的影响分析表明,低压静电场处理能抑制质量损失率和丙二醛含量的上升,减缓软化速度,提高类黄酮和抗坏血酸等抗氧化物质的含量,抑制多酚氧化酶和过氧化物酶活性,从而延缓双孢蘑菇发生褐变。胡欣悦等[21]研究表明,双孢蘑菇经脉冲强光处理后多酚氧化酶活性降低,过氧化氢酶活性升高,表明脉冲强光可以通过抑制酚类物质氧化、提高双孢蘑菇抗氧化能力来延缓双孢蘑菇发生褐变。多酚氧化酶和过氧化物酶协同作用催化酚类、类黄酮的氧化和聚合,二者活性均呈先上升后下降趋势,可能是因为过氧化物酶在褐变前期发挥重要作用,多酚氧化酶在后期发挥重要作用,这与王礼群等[22]的研究结果基本一致。酪氨酸酶活性整体呈下降趋势,最后维持低活性状态;漆酶活性保持较低水平状态,说明二者可能在双孢蘑菇褐变过程中发挥作用较小。
综上所述,在双孢蘑菇褐变过程中,随着水分的不断流失,渗透调节物质发生波动以应对褐变反应,相关酶活性发生变化,多酚氧化酶和过氧化物酶协同推动褐变的进程。