

密点麻蜥(Eremias multiocellata)属有鳞目(Reptilia)蜥蜴亚目(Squamata)蜥蜴科(Lacertidae)麻蜥属(Eremias),是卵胎生变温动物,主要以昆虫为食[1],常见于北部和西北部的荒漠草原和半荒漠草原[2]。该物种不仅在维持生态平衡方面具有重要作用,还是一种具有较大开发潜力的动物药[3]。目前,有关其药用作用的研究越来越多,如应用密点麻蜥治疗肝炎肝纤维化已取得较好疗效,其可有效改善患者症状[4];其具有较好的抑制人胃癌MKN45细胞铁死亡的作用[5],因此加强对野生密点麻蜥种质资源的保护和开发利用具有重要的现实意义。
线粒体DNA作为重要的遗传信息载体之一,具有相对分子量低、结构简单、拷贝数高、母系遗传和无重组等特点,常用于物种鉴定、母系数量、系统发育和起源分析等[6]。线粒体ATP6基因在细胞能量代谢、遗传稳定性、疾病相关性、编辑位点多样性和单拷贝性等方面发挥着重要作用,使细胞能够高效产生能量、准确传递遗传信息、适应不同环境和生理条件,以及维持细胞功能的正常进行。另外,ATP6编码的ATP合酶a亚基对蛋白质结构与作用的研究具有重要价值。线粒体控制区(D-loop)位于tRNAPro 和rRNAPhe 基因之间,不参与编码蛋白,是mtDNA序列中变异较大的区域,进化速度快,适用于亲缘关系较近群体间的比较和种群水平的差异研究[7]。本研究以甘肃、宁夏和内蒙古地区的8个采样区域共计42只密点麻蜥为研究对象,对其线粒体ATP6基因和D-loop区序列进行扩增,然后进行遗传多样性和系统发育分析,以补充西北部分地区的密点麻蜥基础遗传资源,为保护和开发其种质资源提供参考。
1 材料与方法
1.1 试验材料
样本采自甘肃、宁夏和内蒙古的8个区域,共采集了42只密点麻蜥的肌肉样本(表1)。将采集的样本置于无水乙醇与生理盐水(1∶1)制成的组织保存液中,-20 ℃保存,用于后续分子生物学实验。
表1 采样信息 |
| 样本编号 | 采样区域 |
|---|---|
| M22、M23、M24、M25、M26、M27、M28、M29、M30、M31、M32、M33、M34 | A甘肃盐池湾 |
| M37 | B甘肃金昌赵家沟 |
| M44、M45、M46、M47、M48、M49、M50、M51、M52、M53、M54 | C甘肃马繁山 |
| M55、M56、M57、M63、M65、M68、M69、M71、M72、M73 | D宁夏陶乐 |
| M74、M75、M76、M87 | E宁夏沙坡头 |
| M177 | F宁夏甘塘 |
| M96 | G内蒙古鄂前旗 |
| M100 | H内蒙古乌审旗 |
1.2 试验试剂与仪器
DNA提取试剂盒(湖南艾科瑞生物工程有限公司);Premix Taq(Takara);紫外分光光度计、Tanon 1600 凝胶成像仪器(上海天能科技有限公司)。
1.3 基因组提取与扩增
使用DNA提取试剂盒提取样本DNA,并置于-20 ℃保存。使用紫外分光光度计和琼脂糖凝胶电泳测定DNA浓度。根据GenBank中的密点麻蜥线粒体全序列,利用Primer Premier 5.0、Primer-BLAST和DNASTAR软件分别设计2对引物,并参考得分值,选择最优引物(表2)。引物合成由湖南艾科瑞生物工程有限公司完成。
表2 引物序列信息 |
| 引物 | 引物序列(5’-3’) | T m/℃ |
|---|---|---|
| F—ATP6 | ACAGTGGCTAAAGTTGCGGA | 55.1 |
| R—ATP6 | GTTACCCTGGGAGTTCCACT | |
| F—D-loop | TTTAGTGTCGAAAAATATGTCTACCAA | 55.1 |
| R—D-loop | ACATTAACTGATTTACCCCACGAT | |
ATP6基因的PCR扩增体系(25 μL):模板DNA 2 μL,浓度50 ng/μL;上、下游引物各1.5 μL,浓度10 mmol/L;Premix Taq 12 μL,浓度5 U/μL;无菌水8 μL。D-loop区序列的PCR扩增体系(25 μL):模板DNA 2 μL,浓度50 ng/μL;上、下游引物各0.5 μL,浓度10 mmol/L;Premix Taq 12.5 μL,浓度5 U/μL;无菌水9.5 μL。
PCR反应条件:94 ℃预变性3 min;94 ℃变性45 s,55.1 ℃退火45 s,72 ℃延伸1 min,35个循环;最后72 ℃延伸10 min,4 ℃保存。
PCR扩增产物使用1.4%琼脂糖凝胶进行电泳检测,使用Tanon 1600凝胶成像仪器观察获得的PCR扩增产物条带是否达到目标亮度,条带中是否存在引物二聚体和假阳性的情况,并将扩增成功的PCR产物送至测序公司进行测序。
1.4 数据处理与分析
利用Geneious2024.0.5(https://www.geneious.com)对测序结果进行处理,将对应的上下游测序结果序列拼接到一起,经过人工校对得到密点麻蜥ATP6基因和D-loop区序列各42条。从GenBank上下载其他麻蜥属蜥蜴的D-loop和ATP6基因序列,作为外群物种序列(表3),与校对完成的42条密点麻蜥的样本序列进行对比分析,最终确定D-loop序列的长度为712 bp,ATP6基因序列的长度为489 bp。将分析结果以fasta格式导出,做下一步分析。
表3 GenBank下载的麻蜥序列信息 |
| 中文学名 | 拉丁学名 | 序列号 |
|---|---|---|
| 丽斑麻蜥 | Eremias argus | JQ086345.1 |
| 准噶尔麻蜥 | Eremias dzungarica | NC_056302.1 |
| 山地麻蜥 | Eremias brenchleyi | EF490071.1 |
| 快步麻蜥 | Eremias velox | KM359148.1 |
| 天山麻蜥 | Eremias stummeri | NC_029878.1 |
1.4.1 序列分析
采用MEGA-X软件对序列进行多重比较,分析ATP6基因和D-loop区序列的碱基组成、变异位点等信息[8]。
1.4.2 遗传多样性分析
采用DnaSP 6.1.2软件对序列的遗传多样性进行分析,统计序列的单倍型(h),单倍型多样性(Hd)、核苷酸多样性(π)和Tajima’sD值等[9]。
1.4.3 系统发育分析
使用MEGA-X软件,Bootstrap(自展值)设为1 000次,以最大邻接法(NJ)和最大似然法(ML)分别构建NJ树和ML树,在使用NJ法构建进化树中,采用p-distance法计算遗传距离。在使用ML法构建进化树中,首先需要使用Find Best DNA/Protein Models(ML)的功能,找到BIC分数最低的模型作为最佳模型,基于该模型构建ML进化树。通过该功能可以得知,基于ATP6基因构建ML树的最佳模型为TN93(Tamura-Nei)+I,基于D-loop区序列构建ML树的最佳模型为HKY(Hasegawa-Kishino-Yano)+G+I。
2 结果与分析
2.1 PCR扩增
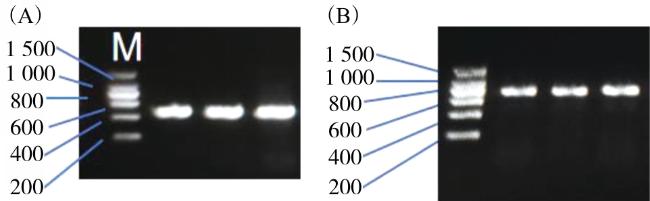
由图1可知,目的条带符合预期大小(扩增的ATP6基因和D-loop区序列目的条带大小分别在400~600 bp和650~800 bp且清晰,条带中不存在引物二聚体和假阳性的干扰,以部分样品在2种扩增体系中的电泳结果为例。
2.2 序列分析
2.2.1 ATP6序列
在ATP6基因序列中A、T、G、C的平均含量分别为26.5%、29.1%、31.9%、12.5%,A+T=55.6%,C+G=44.4%;A+T含量高于G+C含量,碱基组成具有偏倚性。ATP6基因序列中包含保守位点363个,变异位点111个,单态突变位点36个,简约信息位点75个,插入或缺失位点15个。
2.2.2 D-loop区序列
D-loop区序列中A、T、G、C的平均含量分别为34.4%、27.3%、26.1%、12.2%,A+T= 61.7%,C+G=38.3%,A+T含量显著高于G+C含量,碱基组成具有偏倚性。D-loop区序列中包含保守位点618个,变异位点54个,单态突变位点12个,简约信息位点42个,插入或缺失位点40个。
2.3 遗传多样性分析
由于B、F、G和H区域采集的样本数较少,因此仅对A、C、D和E 4个采样区域的密点麻蜥种群进行遗传多样性分析。由表4可知,基于ATP6基因,A点的密点麻蜥单倍型数量(h)为3个,单倍型多样性(Hd)为0.500,核苷酸多样性(π)为0.006 93,平均核苷酸差异(k)为3.307 69,Tajima’s D值为-2.082 26<0。C点的密点麻蜥h为5个,Hd为0.709,π为0.012 12。k为5.781 82,Tajima’s D值为-2.105 80<0。D点的密点麻蜥h为8个,Hd为0.933,π为0.013 78,k为6.533 33,Tajima’s D值为-0.755 88<0。E点的密点麻蜥h为3个,Hd为0.833,π为0.008 39,k为4.000 00,Tajima’s D值为-0.824 07<0。
表4 密点麻蜥遗传多样性参数 |
| 序列 | 采样区域 | 单倍型数量(h)/个 | 单倍型多样性(Hd) | 核苷酸多样性(π) | 平均核苷酸差异(k) | Tajima’s D值 |
|---|---|---|---|---|---|---|
| ATP6 | A | 3 | 0.500 | 0.006 93 | 3.307 69 | -2.082 26 |
| C | 5 | 0.709 | 0.012 12 | 5.781 82 | -2.105 80 | |
| D | 8 | 0.933 | 0.013 78 | 6.533 33 | -0.755 88 | |
| E | 3 | 0.833 | 0.008 39 | 4.000 00 | -0.824 07 | |
| D-loop | A | 4 | 0.603 | 0.002 83 | 1.923 08 | -1.849 23 |
| C | 5 | 0.618 | 0.005 85 | 3.963 64 | -2.032 39 | |
| D | 6 | 0.844 | 0.007 77 | 5.288 89 | 1.114 91 | |
| E | 3 | 0.833 | 0.005 14 | 3.500 00 | -0.817 34 | |
基于D-loop区序列,A点的密点麻蜥h为4个,Hd为0.603,π为0.002 83,k为1.923 08,Tajima’s D值为-1.849 23<0。C点的密点麻蜥h为5个,Hd为0.618,π为0.005 85,k为3.963 64,Tajima’s D值为-2.032 39<0。D点的密点麻蜥h为6个,Hd为0.844,π为0.007 77,k为5.288 89,Tajima’s D值为1.114 91>0。E点的密点麻蜥h为3个,Hd为0.833,π为0.005 14,k为3.500 00,Tajima’s D值为-0.817 34<0。其中,只有D点密点麻蜥的Tajima’s D值大于0,说明其密点麻蜥的单倍型数大于变异位点数,稀有等位基因较少,该地区的密点麻蜥种群可能没有经历过种群扩张。
综合4个采样区域,D点的密点麻蜥ATP6基因和D-loop区序列的h,Hd和π的值均最大,表明该区域的密点麻蜥种群的遗传变异程度最高
2.4 系统发育分析
2.4.1 ATP6基因
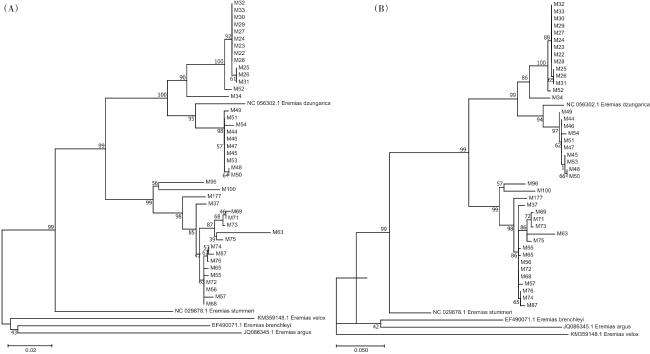
基于ATP6基因构建的NJ树和ML树基本一致,所有密点麻蜥样本位于同一分支上,天山麻蜥、准噶尔麻蜥与密点麻蜥样本聚为一支,说明其亲缘关系较近;尤其是准噶尔麻蜥,与C点的密点麻蜥样本聚为一小支,亲缘关系更为接近。而丽斑麻蜥、山地麻蜥和快步麻蜥聚为另外一支,且丽斑麻蜥和山地麻蜥聚为一小支,三者为明显外群,与样本密点麻蜥的亲缘关系较远。密点麻蜥样本分为明显两支,甘肃地区为一支,宁夏和内蒙古地区为一支。在甘肃地区的密点麻蜥样本中,A点的密点麻蜥样本除M34以外全部位于同一分支上,C点的密点麻蜥样本除了M52以外全部位于同一分支上,亲缘关系密切。而M34与M52与A点的样本聚为一支,表明与其亲缘关系接近。M37(B)则与宁夏地区的密点麻蜥样本聚为一支,与甘肃地区的密点麻蜥样本亲缘关系较远,在内蒙古地区的密点麻蜥样本中,M96(G)和M100(H)聚为一支,亲缘关系密切,并且与宁夏地区的密点麻蜥样本亲缘关系较为接近。在宁夏地区的密点麻蜥样本中,M177(F)按采样区域独立分布,而E和D点的样本均聚为一支,M74、M76和M87(E)聚为一小支,表示亲缘关系更为密切,但M75(E)与D点的部分密点麻蜥样本聚为一小支,表示两者间样本亲缘关系更近。以上聚类结果说明宁夏和内蒙古地区的密点麻蜥之间可能存在频繁交流(图2)。
2.4.2 D-loop区序列
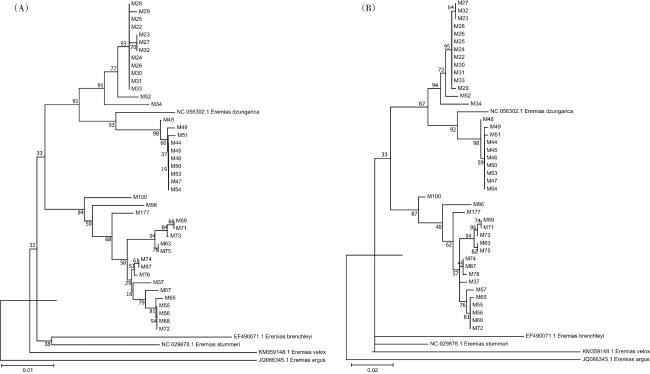
由图3可得,基于D-loop区序列构建的NJ树和ML树基本一致,与ATP6基因相比,样本在不同进化树中的位置和关系也基本相似。有所不同的是,从物种间关系上来看,在基于D-loop区序列构建的NJ树中,丽斑麻蜥和密点麻蜥不在同一分支上,与密点麻蜥亲缘关系较远,其余麻蜥均与密点麻蜥位于同一分支上,并且山地麻蜥和天山麻蜥聚为一小支,亲缘关系更近。而在基于D-loop区序列构建的ML树中,除准噶尔麻蜥与C样本点的密点麻蜥样本聚为一小支外,其余不同麻蜥种间没有聚为一小支。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
本研究中密点麻蜥ATP6基因和D-loop区序列的A+T含量分别为55.6%和61.7%,由于碱基A+T含量高的线粒体基因在进化中更具优势[10],因此相较于ATP6基因,D-loop在研究密点麻蜥的进化关系中更具有优势。ATP6基因和D-loop的Tajima’s D值除陶乐地区的D-loop中密点麻蜥的Tajima’s D值大于0外,其他地区均小于0,表明陶乐地区密点麻蜥的单倍型数大于变异位点的个数,表示稀有等位基因是以低频率存在,推断该地区种群可能未经历过种群扩张。其他地区密点麻蜥的单倍型数小于变异位点的个数,表示稀有等位基因是以高频率存在,推测在其他地区种群中低频等位基因占主导作用。
系统发育分析表明,密点麻蜥样本种群与其他5种麻蜥属麻蜥存在不同程度的亲缘关系,且明显聚为两支,甘肃地区为一支,宁夏和内蒙古地区为一支。在甘肃地区的密点麻蜥样本中,A和C区域之间部分样本亲缘关系较近,说明两地之间可能存在基因交流现象,并且B区域中M37与宁夏地区的密点麻蜥样本聚为一支,表明该地与宁夏地区的密点麻蜥样本间可能存在基因交流。在宁夏地区的密点麻蜥样本中,E和D的样本均聚为一支,表明两地之间存在频繁的基因交流现象。在内蒙古地区的密点麻蜥样本中,虽然M96、M100均与宁夏地区的密点麻蜥聚为一支,但在进化树中的位置距离较远,推测内蒙古地区的密点麻蜥样本与宁夏地区的密点麻蜥样本间可能存在轻度的基因交流。基因交流一方面可以促进遗传变异,影响群体遗传多样性,产生新的性状组合;另一方面,过度的基因交流导致物种间差异减少、种群发生近交衰退和遗传多样性减少等,可能导致原始物种消失,造成优良种质资源不可逆地减少,从而破坏生态平衡。
综上,本研究分析了不同地区密点麻蜥ATP6基因和D-loop区序列的遗传多样性及系统发育关系,结果表明,相比于ATP6基因,D-loop在研究密点麻蜥的进化关系中更具有优势,因此在未来有关密点麻蜥进化关系的研究中可以偏向于D-loop;陶乐地区密点麻蜥样本的遗传变异程度相比其他地区更高,可以进一步探索出现该现象的原因;在甘肃、宁夏和内蒙古地区的密点麻蜥种群间均存在基因交流,过度的基因交流在保护优良的种质资源上是不利的,但在一定程度上有利于增加物种的遗传多样性,因此,需要加强对密点麻蜥的遗传多样性监测。