

盐胁迫是影响植物生长的重要逆境因素之一,高渗透压会使植物细胞失水、气孔关闭、呼吸速率下降,影响生命活动。蒸腾作用导致细胞失水,无机盐离子积累导致细胞渗透压失衡,对植物生长有毒害和抑制作用[1-2]。土地盐碱化是土壤含盐水分蒸发后溶质积累的过程,所引起的盐胁迫易导致作物产量下降[3]。
植物抗氧化酶系统是抵御活性氧毒性、维持稳态和参与环境胁迫信号转导的保护系统,抗氧化酶包括超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化物酶(Peroxidase,POD)、抗坏血酸过氧化物酶(Ascorbate peroxidase,APX)和过氧化氢酶(Catalase,CAT)等[4]。在植物遭受盐胁迫时,SOD可清理细胞质和相关细胞器中的超氧阴离子[5];CAT、POD与SOD协作分解产物;APX存在于叶绿体,能清除H2O2。多种酶共同组成系统,可通过观察抗氧化酶含量分析植物抗逆性[6]。
玉簪(Hosta plantaginea)主要分布于四川、湖北、湖南、江苏、安徽、浙江、福建和广东等温暖潮湿地区,其根有药效。该植物多应用于乔木、灌木植物下层,用作装饰;其对营养和光照需求低,也可种在岩石园、背阴面或修饰林地、河边等贫瘠处。其姿态优美、观赏价值较高,是庭院常用植物之一。该植物叶片颜色多样,可用于园艺设计,能点缀林地。众多优良的玉簪品种正被引种至寒冷地区,为高寒地区引种栽培奠定了基础[7]。谢树章等[8]研究发现,‘金标’玉簪生根最佳培养基为1/2 MS+0.50 mg/L IBA,生根率较高;张琳等[9]研究发现,‘夏香’‘皇家标准’和‘如此甜’等9个品种表现良好,观赏价值较高,适宜在陕西宝鸡地区进行园林推广应用。近年来,土壤盐碱化造成了一定的经济损失,提高盐碱地区的土地利用率对减少损失具有重要作用。本试验以玉簪幼苗为对象,设置不同浓度的盐胁迫,测定其CAT 、SOD 、POD和APX 4种过氧化物酶的活性,为提高玉簪在盐碱地的利用率提供参考。
1 材料与方法
1.1 试验材料与仪器
试验材料为玉簪幼苗;试验试剂包括氯化钠、愈创木酚、过氧化氢、磷酸二氢钾、磷酸二氢钠、甲硫氨酸、氮蓝四唑、核黄素溶液、聚乙烯聚吡咯烷酮和EDTA-Na2溶液等,均为化学纯;试验仪器包括紫外分光光度计、超速离心机、恒温培养箱、离心管、电子天平和冰箱等。
1.2 试验设计
1.3 测定指标及方法
1.3.1 CAT活性
使用紫外分光光度法[12]测定CAT活性。取0.2 g玉簪叶片,液氮速冻,用研钵充分研磨后加4 ℃预冷的2 mL PBS(pH 7.8)。液体转移至离心管,摇匀后4 ℃、10 000 r/min离心10 min。取100 μL上清液煮沸10 min,吸取3 mL CAT反应液加入比色皿,在其中加入80 μL上清液混匀,再加80 μL PBS调零,立刻计时,每隔30 s记录240 nm处的吸光度值5~6次。计算如式(1 )~(2 )。
煮沸后酶液的吸光度值=(30 s时所测的吸光值+3 min时所测的吸光值)/2
CAT活性=煮沸后酶液的吸光度值/(0.1×测定时提取液的用量×反应时间×植物鲜重)
1.3.2 SOD活性
参考程艳等[13]的方法测定SOD活性。分别称取各处理组玉簪去脉叶片0.5 g,在预冷研钵中加入2 mL磷酸缓冲液研磨成匀浆,转移至10 mL容量瓶。取5 mL提取液4 ℃、10 000 r/min离心15 min,取上清液(酶粗提液)。取7支离心管置于冰上,1~3号管为测定管,加入1.5 mL 50 mmol/L磷酸缓冲液,再加0.3 mL 130 mmol/L 甲硫氨酸溶液、0.1 mL粗酶液和0.5 mL蒸馏水;4~7号管为样品管,加入1.5 mL 50 mmol/L磷酸缓冲液和0.6 mL蒸馏水。7号管加试剂后遮光处理,其余6管在常温灯光下显色15~20 min。显色结束,7支试管均避光处理。使用分光光度计在560 nm处测定吸光度值,计算如式(3) 。
SOD活性=(对照管吸光度值-样品管吸光度值)×样品提取液总体积×60/对照管吸光度值×0.5×样品鲜重×测定时粗酶液量×
1.3.3 POD活性
采用愈创木酚比色法[12]测定POD活性。冷却19 μL 30% H2O2,混合后低温保存,配置POD反应液。取0.1 mL磷酸缓冲液(pH 6.8)50 mL、愈创木酚28 μL,80 ℃加热10 min搅拌至愈创木酚溶解,避光冷却,加入19 μL H2O2,混匀后4 ℃避光保存。取0.1 g玉簪叶片,液氮速冻研磨后加4 ℃预冷的2 mL 20 mmol/L磷酸二氢钾溶液,转移至离心管,摇匀后4 ℃、10 000 r/min离心10 min,吸取3 mL POD反应液和160 μL上清液,混匀加进比色皿。用PBS(pH 6.0)代替上清液与POD反应液混匀,用于分光光度计调零,在470 nm处测吸光度值,每隔30 s测定1次,共5次,总计时短于210 s,计算如式(4) 。
POD活性=(末次吸光度值-首次吸光度值)×样品提取液总体积/0.01×植物鲜重×测定时提取液用量×反应时间
1.3.4 APX活性
参考杜佳庚等[14]测定APX活性的方法。将0.5 g幼苗叶片剪碎,在研钵加石英砂等磨料和适量液氮速冻,置于冰上充分研磨,加入7.5 mL提取液与粉末混匀,液体转至离心管,4 ℃、10 000 r/min离心10 min,吸取20 μL上清液和3 mL反应液混匀后分别加入3支离心管,1号管加等体积水作空白对照,2号管加AsA,3号管加0.1 mmol/L H2O2,充分混匀后转至比色皿,置于预热分光光度计测定290 nm处的吸光度值,反应启动后每10 s测定1次,计算如式(5) 。
APX活性=吸光度值×7.5×1 000×60×1 000/植物鲜重×2.8×20×10
1.4 数据分析
采用SPSS和Excel软件进行数据分析。
2 结果与分析
2.1 对玉簪幼苗CAT活性的影响
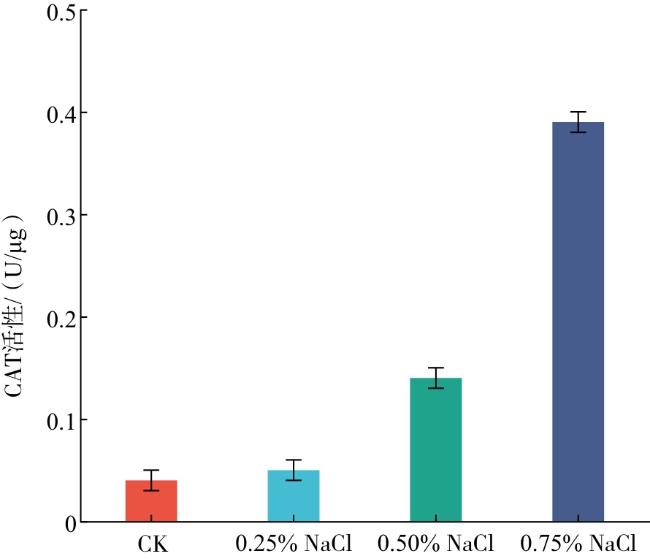
由图1可知,随着NaCl溶液浓度的提高,CAT活性呈上升趋势,其中0.25% NaCl胁迫处理下玉簪幼苗的CAT活性较CK提高了5.03%,说明CAT活性升高有助于清除植物体内多余的H2O2,避免H2O2积累对细胞造成破坏;0.50% NaCl处理下幼苗的CAT活性较CK提高了204.58%,较0.25% NaCl处理提高190.00%;0.75% NaCl溶液处理下幼苗的CAT活性较CK提高了770.22%,较0.50%NaCl处理提高了185.71%。说明在0.75% NaCl胁迫下的玉簪幼苗CAT活性较强。
2.2 对玉簪幼苗SOD活性的影响
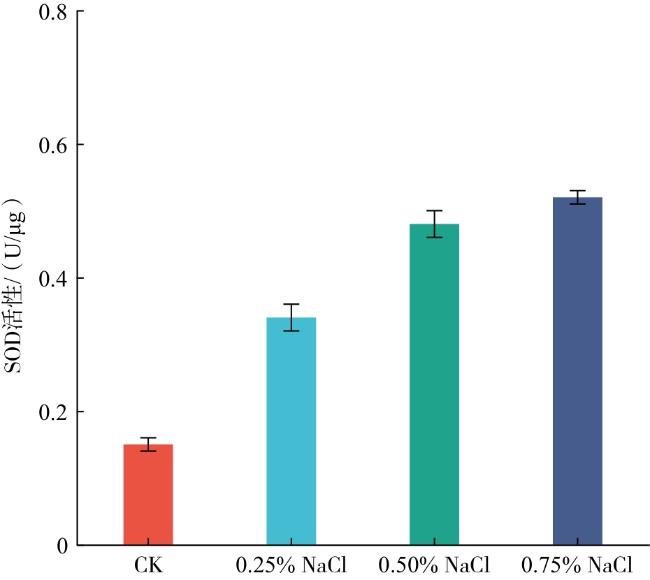
由图2可知,随着NaCl溶液浓度的提高,玉簪幼苗的SOD活性呈上升趋势。其中0.25% NaCl处理下幼苗的SOD活性较CK提高117.46%;0.50% NaCl处理下幼苗的SOD活性较CK提高了208.32%,较0.25% NaCl处理提高了41.78%;0.75% NaCl处理下幼苗的POD活性较CK提高了238.06%,较0.50% NaCl处理提高了9.65%。表明SOD在玉簪抵御盐胁迫过程中发挥重要作用,其对0.75%NaCl胁迫的响应较强。
2.3 对玉簪幼苗POD活性的影响
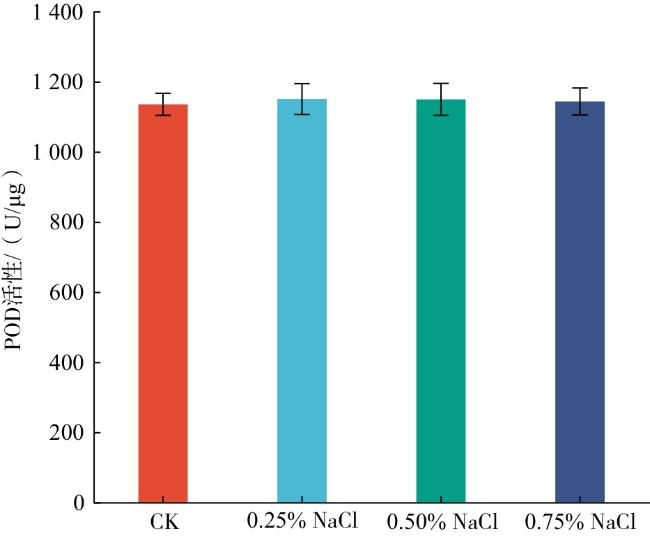
由图3可知,0.25% NaCl处理下叶片的POD活性较CK提升1.32%。这表明POD可能与SOD、CAT共同参与清除玉簪幼苗体内过剩的自由基。0.50% NaCl处理下叶片的POD活性较CK提升1.24%,较0.25% NaCl处理下降0.08%;0.75% NaCl处理下叶片的POD活性较CK上升0.72%,较0.50% NaCl处理下叶片的POD活性下降0.51%,说明POD对0.50% NaCl胁迫响应较强。
2.4 对玉簪幼苗APX活性的影响
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
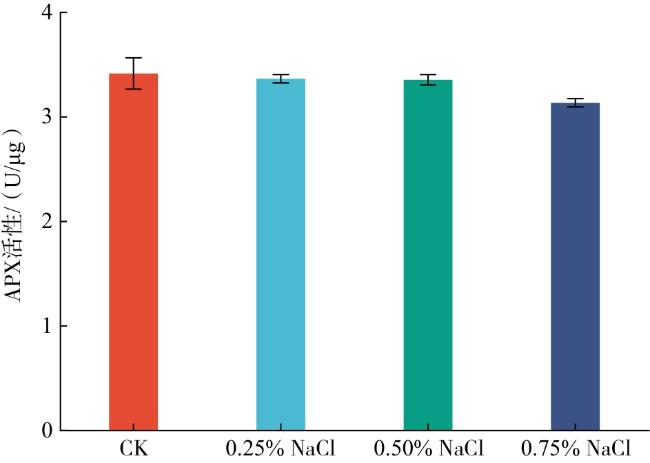
当玉簪幼苗处于盐胁迫环境时,叶片会产生一系列生理应答机制,具体表现为通过提高叶片中的相关酶活性,增强植株对叶片中异常积累的强氧化性自由基以及活性氧基团的清除能力,从而减轻细胞受到的伤害。里程辉等[15]研究发现,在植物淹水过程中,各砧穗组合叶片和根系中SOD、POD、CAT和APX活性大体呈先上升后下降趋势,可减轻对植物细胞的损伤。本试验结果表明,随着NaCl溶液浓度的提高,玉簪叶片CAT、SOD和POD活性大体呈上升趋势,其中SOD活性出现上升速率逐渐缓慢的趋势,与孙方行等[16]研究结果相似,可能是玉簪通过提高SOD活性清除细胞内产生的活性氧自由基的能力有限,当外界NaCl浓度过高、超出其承受阈值时,细胞自由基的清除量小于产生量,易对细胞造成伤害。当盐浓度由0.25%提升至0.50%时,玉簪POD活性呈现缓慢下降趋势。随着NaCl溶液浓度的提高,玉簪叶片APX活性呈现下降趋势,与王立凤等[17]研究结果不同,可能是北五味子属耐盐植物,承受盐浓度阈值比玉簪高。
综上,本研究表明,当盐胁迫浓度达0.25%时,CAT、POD和APX活性与对照相比变化较小,说明低浓度盐胁迫下,玉簪幼苗CAT、POD和APX响应盐胁迫的程度较小。当盐胁迫浓度达0.50%时,CAT、SOD和POD活性较对照组升高;盐浓度达0.75%时,CAT、SOD和POD活性继续缓慢提升,APX活性则持续下降。随着盐浓度的提高,玉簪幼苗CAT、SOD和POD 3种抗氧化酶活性大体呈上升趋势,APX则呈下降趋势。说明玉簪幼苗主要通过提高CAT、SOD和POD活性、降低APX活性提高调节抗氧化系统,以抵御盐胁迫。