

金钗石斛(Dendrobium nobile)是兰科石斛属的 多年生草本植物,是药用石斛的主要来源品种之一[1]。该植物含生物碱、倍半萜、菲和联苄类等,具有滋阴、清热、润肺、止咳和明目等功效[2]。转录组学分析在植物发育和胁迫应答机制等领域应用广泛[3]。Domingos等[4]利用转录组学对葡萄花进行测序,识别发育关键基因。相关研究还有铁皮石斛花[5]、锁阳花[6]、灰毡毛忍冬花[7]、威氏绿绒蒿花[8]、印度菊花[9]、木棉花[10]和茶树花[11]等。低温、高温和机械损伤是常见的非生物胁迫,通过转录组学成功识别了拟南芥[12]、马铃薯[13]、番茄[14]和高山离子芥[15]等多种植物冷应答基因。Li等[16]分析了柳枝稷高温应答基因。利用测序,识别了白木香[17]、樟树[18]和土沉香[19]等植物的机械损伤应答基因。
MADS是一类重要的转录因子,是调控植物生长、发育和生殖等过程的主要调控因子之一,尤其是在植物花、果实、根、茎、叶发育等过程中起到重要作用[20]。相关学者利用分子生物学技术,对石斛属植物MADS转录因子编码基因进行克隆与功能分析。例如,袁慧等[21]采用PCR和RACE技术对铁皮石斛AP1-A MADS-box和AP1-B MADS-box进行克隆,并对基因进行表达分析。Chen等[22]鉴定了16个铁皮石斛MADS基因,其中8个为特异性表达。金钗石斛在生药学、药理和化学等方面已有深入研究,但在发育和胁迫应答关键基因挖掘等生物技术领域的研究较少。因此,本研究利用转录组学,对金钗石斛花发育和胁迫的转录模式和MADS差异表达开展分析,为解析分子机制提供参考。
1 材料与方法
1.1 试验材料
金钗石斛植株由贵州中医药大学生药学实验室提供,选择小花苞期(呈黄绿色)、大花苞期(呈部分紫色,花瓣未展开)和开放期(呈紫色,花瓣完全展开)作为花的3个发育时期(Ⅰ期、Ⅱ期、Ⅲ期)。使用开放期同类型材料进行低温(4 ℃,1 h)、高温(40 ℃,1 h)和机械损伤(将材料剪出间隔1 cm,深度0.5 cm的伤口,1 h)3种非生物胁迫。收集样本,等量混样后,液氮冷冻保存,用于后续试验。将3个发育时期样本进行Ⅰ期vsⅡ期、Ⅱ期vsⅢ期、Ⅰ期vsⅢ期分组对比,将3种非生物胁迫分别与无胁迫的Ⅲ期样本进行对比。
1.2 试验方法
利用华大基因Illumina Ⅱ系统进行测序,利用SOAPnuke v1.5.2软件对原始数据进行过滤,包括删除“污染”读长(Reads)、未知碱基超过10%的读长、低质量碱基超过50%的读长,以FASTQ格式存储。然后,利用Trinity v2.0.6软件从头组装,BUSCO软件检测组装质量,产生单基因(Unigene)。利用Transdecoder v3.0.1预测编码区;利用Nr、Nt、SwissProt、KEGG、COG、Pfam和GO数据库注释。利用RSEM v1.2.8和DESeq计算Unigene表达量及组内差异,以大于2倍且P value≤0.001为显著差异。差异表达基因(DEGs)根据GO和KEGG等数据库进行注释和分类,以Q value≤0.05为显著标准。基于测序数据,利用基因注释信息和序列比对,识别MADS转录因子[22]。
2 结果与分析
2.1 差异表达基因
对金钗石斛花3个发育阶段(Ⅰ期、Ⅱ期、Ⅲ期)和3种胁迫(低温、高温和机械损伤)样本进行测序,分别获得42.1 M、42.26 M、43.25 M、42.77 M、43.28 M和42.03 M个读长(Reads),Q20均大于98.06%,Q30均大于94.09%。利用从头组装获得155 004个单基因。对Nr、Nt、SwissProt、KEGG、COG、Pfam和GO数据库注释进行分析,发现分别有88 975、93 599、44 832、54 385、41 294、44 074和50 154(共108 566)个单基因获得注释。由图1可知,Ⅰ期vsⅡ期、Ⅱ期vs Ⅲ期和Ⅰ期vsⅢ期分别识别了26 598、29 680和30 263个DEGs;低温、高温和机械损伤分别识别了7 495、13 713和8 764个DEGs。
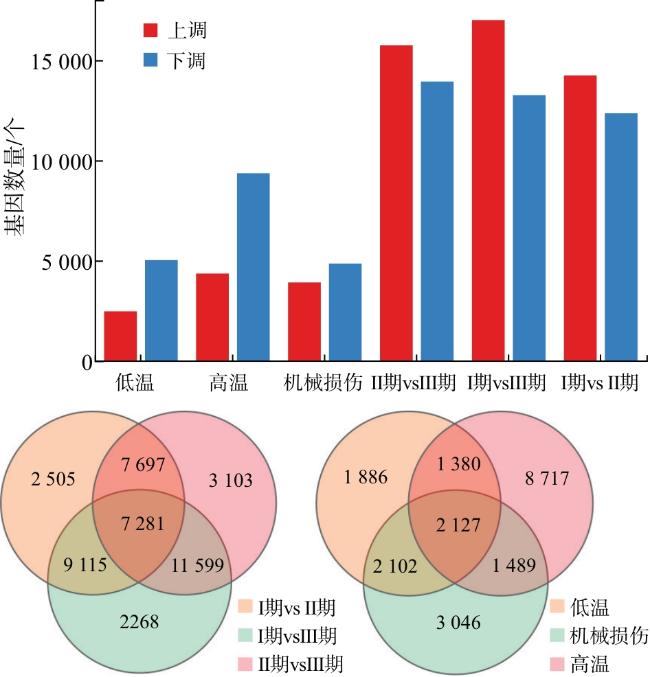
2.2 DEGs的功能聚类和调控通路
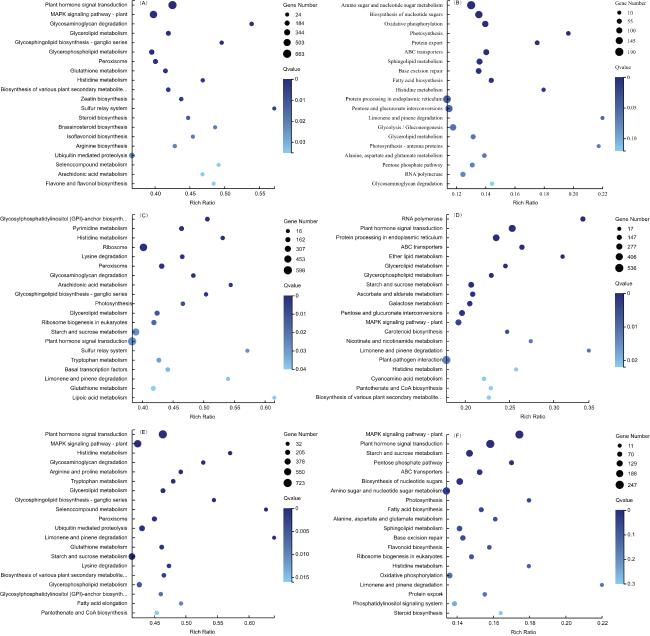
对DEGs进行GO功能聚类,表1列举了每种类别中富集程度最大的2类。结果表明,生物过程、细胞组分和分子功能均发现大量基因聚类,花发育不同阶段之间聚类基因较多,高温胁迫也对基因功能聚类产生较大影响。同时对DEGs进行KEGG调控通路预测(图2),结果表明,花发育不同阶段差异表达基因与植物激素信号转导(Plant hormone signal transduction)、植物MAPK信号通路(MAPK signaling pathway-plant )、淀粉和蔗糖代谢(Starch and sucrose metabolism)等过程有关(图2A、C和E);低温胁迫与氨基糖和核苷酸糖代谢(Amino sugar and nucleotide sugar metabolism)、内质网中的蛋白质加工(Protein processing in endoplasmic reticulum)等过程有关(图2B);高温胁迫与植物病原体相互作用(Plant-pathogen interaction)等过程有关(图2D);机械损伤与植物MAPK信号通路等过程有关(图2F)。功能聚类和调控通路分析为宏观层面揭示金钗石斛分子机制提供基础,显示了基因的类群与功能细分。
表1 差异表达基因的GO聚类单位:个 |
| 样本 | 生物过程 | 细胞组分 | 分子功能 | |||
|---|---|---|---|---|---|---|
| 细胞过程 | 代谢过程 | 细胞解剖实体 | 蛋白复合物 | 催化活性 | 结合 | |
| Ⅰ期vsⅡ期 | 5 092 | 4 734 | 7 633 | 857 | 7 055 | 6 931 |
| Ⅱ期vsⅢ期 | 5 820 | 5 290 | 8 269 | 1 085 | 7 582 | 7 621 |
| Ⅰ期vsⅢ期 | 5 675 | 5 242 | 8 572 | 953 | 7 885 | 7 743 |
| 低温 | 1 529 | 1 375 | 1 918 | 238 | 1 852 | 2 208 |
| 高温 | 2 463 | 2 202 | 3 589 | 295 | 3 293 | 3 842 |
| 机械损伤 | 1 688 | 1 520 | 2 373 | 244 | 2 198 | 2 560 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.3 MADS转录因子的差异表达分析
转录因子与真核基因启动子区域的顺式作用元件互作,调控基因表达。金钗石斛无基因组序列,基于转录组水平的测序为挖掘基因功能提供了一种可行的方式。基于测序数据,识别了74个MADS转录因子,其中Ⅰ期vsⅡ期有18个上调基因和22个下调基因、Ⅱ期vsⅢ期有22个上调基因和12个下调基因、Ⅰ期vsⅢ期有23个上调基因和17个下调基因;低温有7个上调基因和10个下调基因、高温有7个上调基因和18个下调基因、机械损伤有4个上调基因和12个下调基因,说明金钗石斛MADS可能在花器官发育和胁迫应答中起到调控作用。表2列举了部分MADS转录因子,DN10042_c0_g1_i1-55A的Ⅰ期vsⅡ期、Ⅱ期vsⅢ期和Ⅰ期vsⅢ期、低温、高温和机械损伤样本的FPKM分别为-4.86、9.76、4.90、-2.99、-9.76和-5.95,分别为下调、上调、上调、下调(抑制)、下调(抑制)和下调(抑制)基因。
表2 MADS家族差异表达 |
| 基因编号 | 基因注释 | logFC | |||||
|---|---|---|---|---|---|---|---|
| I期vs II期 | II期vs III期 | I期vs III期 | 低温 | 高温 | 机械损伤 | ||
| DN10042_c0_g1_i1-55A | MADS2 | -4.86 | 9.76 | 4.9 | -2.99 | -9.76 | -5.95 |
| DN1068_c0_g3_i1-55A | MADS21 | 8.47 | 8.47 | -2.03 | -8.47 | -8.47 | |
| DN1068_c1_g1_i1-55A | AGL6 | 2.90 | 3.08 | 5.98 | |||
| DN1074_c1_g1_i1-55A | MADS16 | 12.26 | 12.26 | ||||
| DN1074_c1_g1_i13-55A | MADS16 | 6.81 | 6.10 | -2.23 | |||
| DN1074_c1_g1_i14-55A | MADS16 | 8.56 | 8.56 | -2.09 | |||
| DN1074_c1_g1_i4-55A | MADS16 | 10.2 | 9.58 | -2.11 | |||
| DN1074_c1_g1_i5-55A | MADS16 | -4.09 | 10.17 | 6.08 | |||
| DN11957_c0_g1_i1-57A | CAULIFLOWER A转录变体X1 | -7.08 | 4.81 | -2.27 | |||
| DN11957_c0_g1_i2-57A | CAULIFLOWER A转录变体X1 | -2.16 | |||||
| DN1256_c0_g1_i6-60A | MADS50转录变体X6 | 2.29 | |||||
| DN1256_c0_g1_i9-60A | MADS50转录变体X6 | 2.27 | 3.24 | ||||
| DN15170_c0_g1_i1-55A | MADS AGL29部分 | -2.73 | |||||
| DN1559_c0_g4_i1-59A | MADS2转录变体X1 | 4.76 | 4.05 | ||||
| DN1624_c0_g6_i1-56A | MADS16 | 2.34 | -2.29 | ||||
| DN16321_c2_g1_i1-55A | MADS JOINTLESS转录变体X1 | -4.77 | 3.78 | ||||
| DN1867_c0_g1_i1-55A | MADS box AP3-类似蛋白A | 3.15 | 3.30 | 6.45 | |||
| DN1867_c0_g1_i2-55A | MADS16类似 | 9.68 | 9.68 | ||||
| DN1867_c0_g2_i1-55A | MADS16 | 2.40 | |||||
| DN1867_c0_g2_i2-55A | MADS16 | 9.87 | 9.87 | -2.64 | |||
| DN19438_c0_g2_i1-60A | MADS SVP转录变体X2 | 12.78 | |||||
| DN22662_c0_g1_i1-59A | MADS SVP转录变体X3 | 3.45 | 2.14 | -7.69 | |||
| DN23575_c0_g1_i3-57A | MADS AGL29部分 | -3.08 | -4.75 | -7.83 | 4.75 | ||
| DN23575_c0_g1_i4-57A | MADS AGL29部分 | -7.11 | 5.98 | -5.98 | |||
| DN25403_c0_g1_i1-60A | CAULIFLOWER A转录变体X2 | 11.99 | -11.99 | ||||
| DN25676_c0_g1_i1-59A | MADS32 | 7.51 | -2.70 | 4.81 | 3.05 | ||
| DN2791_c0_g1_i3-56A | CAULIFLOWER A | -2.05 | -3.53 | -4.58 | -4.58 | -4.58 | |
| DN28168_c0_g1_i1-58A | MADS SVP转录变体X2 | 2.35 | |||||
| DN2980_c0_g3_i3-55A | MADS SOC1 | -2.34 | |||||
| DN3067_c0_g3_i1-57A | MADS JOINTLESS转录变体X1 | -9.85 | 5.04 | -4.80 | -5.04 | ||
| DN3067_c0_g3_i2-57A | MADS JOINTLESS转录变体X1 | -6.73 | -6.16 | ||||
| DN3262_c0_g2_i1-59A | MADS16 | 3.30 | -3.39 | ||||
| DN3262_c0_g2_i2-59A | MADS16 | 2.64 | 3.91 | -2.08 | -2.03 | ||
| DN353_c0_g1_i12-56A | MADS16 | 7.55 | -12.9 | -5.36 | 6.19 | 4.46 | 3.32 |
| DN353_c0_g1_i13-56A | MADS16 | 9.91 | -9.91 | 5.95 | 3.91 | 5.78 | |
| DN353_c0_g1_i3-56A | MADS16 | 10.45 | -10.45 | 3.91 | 4.58 | ||
| DN353_c0_g1_i4-56A | MADS16 | 5.98 | -5.98 | 3.46 | |||
| DN353_c0_g1_i9-56A | APETALA3-类似蛋白3_2 | 8.92 | 7.94 | -7.94 | |||
| DN37239_c0_g1_i1-56A | CAULIFLOWER A-类似转录变体X2 | 3.92 | 3.60 | ||||
| DN432_c0_g2_i2-57A | MADS16 | 2.90 | -2.32 | -2.04 | -2.58 | ||
| DN4759_c0_g1_i1-57A | CAULIFLOWER A | -3.08 | -2.16 | -5.23 | |||
| DN4759_c0_g1_i2-57A | CAULIFLOWER A | -3.40 | -4.72 | -4.32 | -4.32 | -4.32 | |
| DN4759_c0_g1_i3-57A | CAULIFLOWER A | -9.73 | -9.73 | ||||
| DN4987_c0_g1_i2-59A | MADS EJ2 | 2.26 | |||||
| DN5334_c0_g1_i2-57A | MADS3类似 | -9.98 | 3.70 | -6.28 | 2.96 | -3.70 | -3.70 |
| DN5334_c0_g1_i3-57A | AGAMOUS-类似蛋白 | -7.06 | 4.58 | -2.48 | -3.68 | -7.58 | -3.68 |
| DN5334_c0_g2_i1-57A | AGAMOUS类似 | -8.64 | -8.64 | 7.10 | 5.49 | ||
| DN5334_c0_g2_i2-57A | AGAMOUS类似 | -7.89 | 5.32 | -2.57 | -5.32 | -5.32 | |
| DN5338_c0_g1_i1-60A | MADS SOC1 | -3.63 | |||||
| DN5338_c0_g2_i3-60A | MADS SOC1 | -2.62 | -2.38 | ||||
| DN5352_c1_g3_i1-55A | MADS CMB1 | 2.32 | |||||
| DN556_c0_g1_i3-60A | MADS16 | 6.02 | 4.49 | -2.45 | |||
| DN556_c0_g1_i5-60A | MADS16 | 7.92 | 8.96 | ||||
| DN5588_c0_g1_i1-57A | MADS6 | -3.62 | -2.62 | -3.35 | |||
| DN578_c0_g1_i4-57A | MADS AGL9同源 | -7.27 | -6.62 | ||||
| DN6272_c0_g1_i2-58A | MADS EJ2 | -2.62 | 2.52 | ||||
| DN7049_c0_g2_i1-57A | MADS AGL65 | -3.81 | -5.23 | ||||
| DN8879_c0_g1_i1-59A | MADS类似 | 7.43 | |||||
| DN9010_c0_g3_i1-56A | MADS AGL9同源 | -2.91 | -2.04 | ||||
| DN9010_c0_g3_i3-56A | MADS AGL9同源 | -2.64 | |||||
| DN9157_c0_g2_i2-58A | MADS3类似 | -8.77 | 6.87 | 2.15 | -6.87 | ||
3 结论与讨论
植物器官生长发育和抗逆应答是复杂的生物过程,受多种类型基因共同调控。随着测序技术在相关领域的深入研究和应用,这些生物学现象涉及的关键基因逐渐显现。He等[5]对铁皮石斛早期花蕾、中期花蕾以及开放花蕾进行测序分析,共识别8 019个差异表达基因和239个分化相关代谢产物,鉴定了参与花发育的MIKC型MADS基因和参与叶绿素、类胡萝卜素等生物合成的数个基因。MADS家族在植物花器官生长发育中起到重要作用[21]。本研究识别了74个金钗石斛MADS转录因子,在花器官不同发育阶段均存在大量差异表达的MADS转录因子,其中Ⅰ期vsⅡ期、Ⅱ期vsⅢ期、Ⅰ期vsⅢ期分别有40、34和40个差异表达的MADS转录因子;低温、高温、机械损伤有17、25、16个差异表达的MADS转录因子,说明金钗石斛MADS在花器官发育和胁迫应答中发挥重要的调控作用。
综上,本研究利用转录组学,对金钗石斛花3个发育阶段(Ⅰ期、Ⅱ期和Ⅲ期)和3种胁迫(低温、高温和机械损伤)开展转录模式及MADS家族表达分析,结果表明,不同发育阶段和非生物胁迫的DEGs主要与生物过程、细胞组分和分子功能等密切相关;识别了74个金钗石斛MADS转录因子,说明金钗石斛MADS在花器官发育和胁迫应答中具有重要作用。研究结果为金钗石斛为代表的石斛属植物的相关分子机制研究提供参考。