

运城盐湖位于山西省运城市南郊(110°52′—111°00′ E,34°35′—35°49′ N),是硫酸钠型内陆湖[1]。该湖泊是重要的食盐产地,目前主要生产芒硝和硫酸镁[2]。因采用“垦畦浇晒法”产盐,盐湖被分成了大小不等的区域,每个区域的湖水盐度有所不同,且湖水盐浓度变化较大,这会对其中微生物的生态群落分布及多样性造成影响[3]。采盐、采硝等活动在盐湖东部区域持续进行,导致其景观与生态环境同其他盐湖相比大不相同。因此,运城盐湖生态体系可能孕育独特的微生物类群,其中的微生物可以适应高盐环境,同时具有耐低温、耐高温、抗辐射和耐有机溶剂等特点,其研究和开发价值较大[4]。
黑泥是一种湖底沉积物,是运城盐湖的独特资源。目前对运城盐湖的相关研究主要集中在污染物分析[5]、藻类植物[6-7]、卤虫[8-9]等方面,但关于黑泥微生物资源暂无深入和系统地研究。Zhang等[10]从运城盐湖黑泥中分离出一株新型中度嗜盐菌C176T,经鉴定为螺杆菌属的一个新种。Su等[11]分离并鉴定了一株耐盐放线菌CFH 90308(T),发现其次生代谢产物对枯草芽孢杆菌、丁香假单胞菌、嗜水气单胞菌和抗药性金黄色葡萄球菌均具有抑菌活性。Cui等[12]分别从运城盐湖沉积物和新疆塔里木盆地盐渍土中分离得到4株嗜盐古菌YCN1(T)、YCN58(T)、LT38(T)和LT62(T),并通过表型特征、系统发育属、系统基因组属和比较基因组分析,进行菌株分类鉴定。Guo等[13]从运城盐湖沉积物中成功分离出了首个新属级物种,并将其命名为Yunchengibacter salinarum YC-2023-2T。Zeng等[14]利用宏基因组学技术对运城盐湖盐渍土壤中嗜盐细菌的多样性进行了分析;李坤珺等[15]采用克隆文库技术对运城盐湖水和淤泥混合样品的嗜盐细菌菌群组成进行了研究。
1 材料与方法
1.1 材料与试剂
1.1.1 样品采集
黑泥样品采自山西运城盐湖2号滩(110°59′ E,34°55′ N),采集表层2 m以下的沉积物,每个处理3个重复,去除杂质后充分混合,晾干后用粉碎机粉碎,再通过旋转气流分级机分选至200~1 000目。将每份黑泥样品分为3个部分,一部分不作处理,记为未灭菌黑泥(BM0);第二份经高压蒸汽灭菌处理,置于高压灭菌锅中,于121 °C灭菌20 min,记为热压灭菌泥(BM-ASS);第三份经Co-60辐照灭菌处理,辐射剂量为4 kGy,辐照时间为12 h,记为辐照灭菌泥(BM-CIS)。
1.1.2 主要试剂与仪器设备
DSX-24L高压灭菌锅(上海申安医疗器械厂);DNA提取试剂盒(上海赛百盛公司);PCR引物(上海美吉生物医药科技有限公司);AxyPrep DNA凝胶回收试剂盒(Axygen公司);TransStart FastPfu DNA聚合酶(北京全式金生物公司);酶标仪(MD公司);QuantiFluor ST蓝色荧光定量系统(Promega公司);MiSeq PE300测序平台(Illumina公司);GeneAmp 9700 PCR仪(ABI公司)。
1.2 试验方法
1.2.1 DNA提取和PCR扩增
使用DNA提取试剂盒进行DNA提取,使用1%琼脂糖凝胶电泳检测提取DNA的质量,使用超微量分光光度计测定DNA的浓度和纯度。细菌16S rRNA基因V3~V4高变区,使用通用引物对338F(5′-CTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTAGHVGGGTWTCTAAT-3′)进行PCR扩增,随后进行高通量测序[20];古菌16S rRNA基因V3~V5区域,使用通用引物对Arch-344F(5′-ACGGGGYGCAGCAGGCGGGA-3′)和Arch-915R(5′-GTGCTCCCCCGCCAATTCCT-3′)进行PCR扩增,随后进行高通量测序[21]。测序由上海美吉生物医药科技有限公司完成。
1.2.2 测序结果质量分析
在上海美吉生物医药科技有限公司的云平台完成微生物交互分析。利用Uparse软件对测序数据进行聚类,对获得的序列以97%的一致性聚类划分为操作分类单元(Operational taxonomic unit,OTU)[22]。
1.2.3 微生物Alpha多样性分析
选择97%相似度的OTU,利用mothur计算不同随机抽样下的α多样性指数,绘制稀释曲线图。利用Mothur软件计算Alpha多样性指数,包括Chao1指数、Simpson指数和Shannon指数,其中Chao1指数可以反映群落丰富度,其数值越大表示群落丰富度越高;Shannon指数和Simpson指数则反映群落多样性程度,Shannon指数值越大表示群落的多样性越高,而Simpson指数值则相反[23]。
1.2.4 细菌及古菌群落组成分析
利用RDP classifier比对Silva 16S rRNA基因数据库(v138)进行OTU物种分类学注释,置信度阈值为70%,并在不同物种分类水平下统计每个样本的群落组成
1.2.5 微生物群落结构的主成分分析
利用R语言中vegan软件包进行主成分分析(PCA)。
2 结果与分析
2.1 测序结果质量
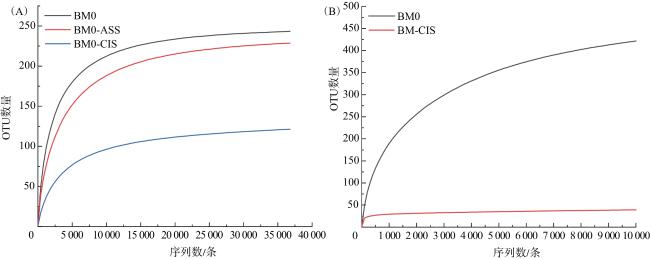
3种黑泥样品测序共得到392 728条细菌优化序列,其中95.62%的样本序列长度在421~440 bp,未灭菌黑泥(BM0)、热压灭菌泥(BM-ASS)和辐照灭菌泥(BM-CIS)中细菌优化序列分别为124 387、131 731和136 610条,覆盖率均大于99%。表明3种黑泥样品的测序数据量足够,可以满足后续分析要求。
3种黑泥样品测序共得到367 779条古菌优化序列,其中长度在261~280 bp的序列占58.63%,241~260 bp的序列占23.65%。BM0和BM-CIS中古菌优化序列分别为104 855和262 924条,而BM-ASS中未测得古菌序列。表明未灭菌黑泥及辐照灭菌泥的测序数据量足够,可以满足后续分析要求;并且热压灭菌黑泥中不含有古菌。
3种黑泥样品在门、纲、属各分类水平上注释物种数如表1所示,从BM0中共检测出细菌29门76纲363属,古菌8门9纲50属,未灭菌黑泥具有丰富的微生物资源。经过灭菌处理后,两种泥样中细菌和古菌的总OTU数及各分类水平上注释物种数均明显减少,物种丰富度明显降低,其中BM-ASS中古菌被完全灭除。
表1 黑泥样品在各分类水平上注释物种数比较 |
| 微生物分类 | 样品 | 总OTU | 门 | 纲 | 属 |
|---|---|---|---|---|---|
| 细菌 | BM0 | 570 | 29 | 76 | 363 |
| BM-ASS | 528 | 22 | 52 | 273 | |
| BM-CIS | 261 | 15 | 42 | 190 | |
| 古菌 | BM0 | 482 | 8 | 9 | 50 |
| BM-ASS | 0 | 0 | 0 | 0 | |
| BM-CIS | 87 | 3 | 4 | 22 |
2.2 微生物Alpha多样性
2.2.1 稀释曲线
2.2.2 多样性指数
3种黑泥样品中细菌和古菌的Alpha多样性指数如表2所示。Chao1值越大说明群落丰富度越高,Shannon值越大、Simpson值越小则说明群落内多样性越高。通常Shannon指数对群落的丰富度以及稀有OTU更敏感,而Simpson指数对均匀度和群落中的优势OTU更敏感[24]。由表2可知,不同灭菌方式处理的黑泥微生物群落多样性存在明显差异。与BM0相比,BM-ASS和BM-CIS的细菌群落Chao1和Shannon指数值均降低,且Simpson指数值均升高。说明经过灭菌处理后,黑泥细菌群落丰富度和多样性均大幅度减少,且Co-60辐照灭菌的减少程度更高。其细菌群落丰富度和多样性从高到低依次为BM0>BM-ASS>BM-CIS。对于古菌群落来说,BM0的Chao1、Shannon和Simpson指数值分别是BM-CIS的5.28倍、1.41倍和1.24倍,说明经过Co-60辐照灭菌处理后,黑泥古菌群落丰富度和多样性均明显减少,而均匀度增加。
表2 各黑泥样品微生物群落多样性指数 |
| 微生物 分类 | 样品 | Chao1指数 | Shannon指数 | Simpson指数 |
|---|---|---|---|---|
| 细菌 | BM0 | 261.0±90.5 | 1.58±0.47 | 0.560±0.120 |
| BM-ASS | 233.0±136.4 | 1.42±0.96 | 0.594±0.263 | |
| BM-CIS | 131.0±7.9 | 0.57±0.09 | 0.829±0.019 | |
| 古菌 | BM0 | 475.0±18.4 | 4.09±0.03 | 0.083±0.005 |
| BM-CIS | 90.0±38.5 | 2.90±0.03 | 0.067±0.002 |
2.3 黑泥细菌群落组成
2.3.1 门水平组成
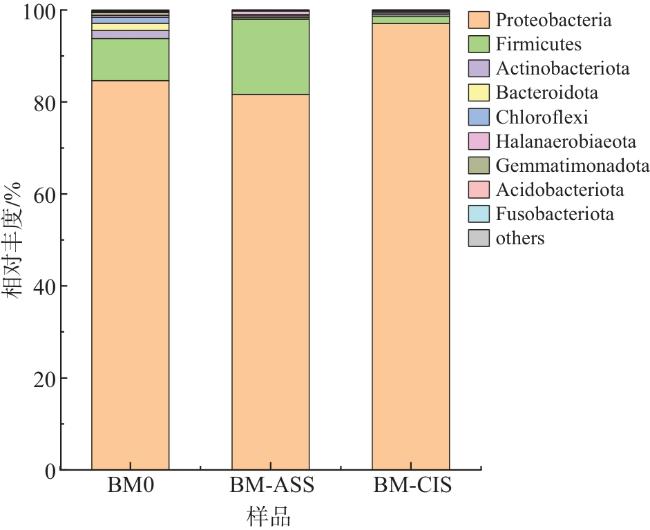
在门水平上,3种黑泥样品中分别检出29、22和15个细菌门(表1)。进一步分析比较门水平下的物种相对丰度可知(图2),3种黑泥样品的优势菌门组成差异不明显,均以变形菌门(Proteobacteria)为优势菌门。在BM0、BM-ASS和BM-CIS中所检出的变形菌门平均相对丰度分别为84.66%、81.68%和97.07%。第二大优势菌门为厚壁菌门(Firmicutes),在BM0、BM-ASS和BM-CIS中的平均相对丰度值分别为9.16%、16.33%和1.56%。与BM0相比,BM-ASS中变形菌门的相对丰度降低,厚壁菌门的相对丰度升高,而BM-CIS中的情况与之相反,变形菌门的相对丰度增加,厚壁菌门的相对丰度降低。说明不同灭菌方式对黑泥中不同细菌丰度的影响不同。
2.3.2 属水平组成
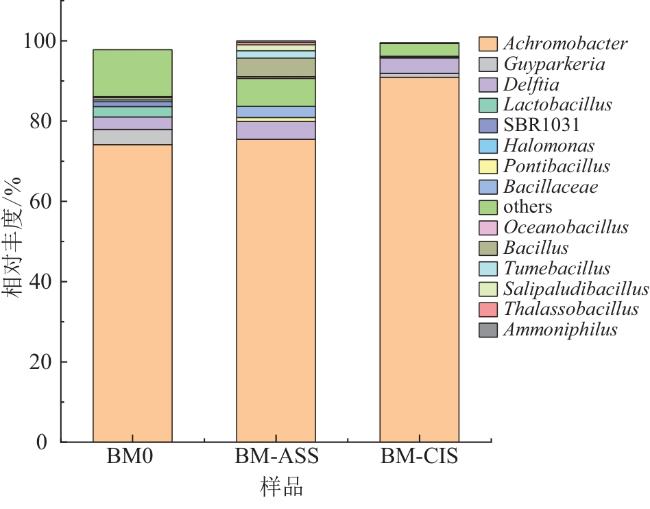
3种黑泥样品中分别检出363个、273个和190个细菌属。对其属水平下的物种相对丰度进一步分析比较可知(图3),3种黑泥样品的优势菌属均为无色杆菌属(Achromobacter),在BM0、BM-ASS和BM-CIS中的平均相对丰度值分别为74.14%、75.46%和90.94%。说明经过灭菌处理后,优势菌的相对丰度值均有提高,但提高幅度有明显区别。代尔夫特菌属(Delftia)的情况类似,在BM0、BM-ASS和BM-CIS中的平均相对丰度值分别为3.12%、4.48%和3.85%。盖伊帕克氏菌属(Guyparkeria)和SBR1031属在BM0中的相对丰度分别为3.78%和1.20%,在BM-ASS中被完全灭除,在BM-CIS中的相对丰度值明显降低,分别为0.94%和0.29%。乳杆菌属(Lactobacillus)和盐单胞菌属(Halomanas)在BM0中的相对丰度分别为2.58%和0.51%,经两种灭菌处理后被完全灭除。
2.4 黑泥古菌群落组成
2.4.1 门水平组成
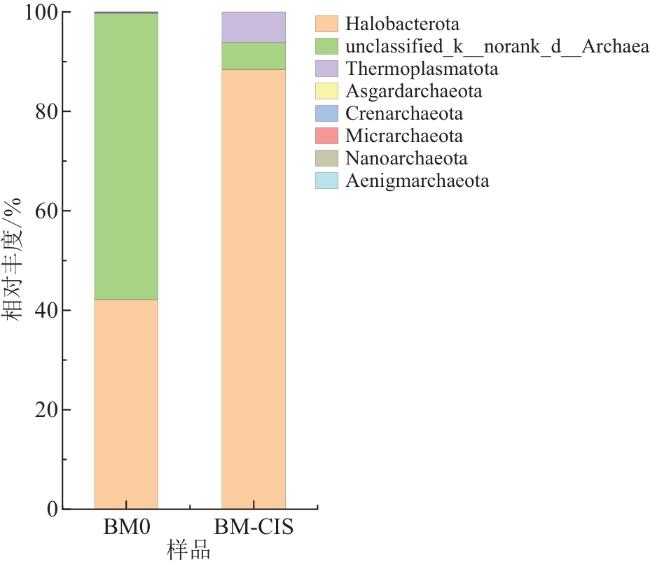
在门水平上,未灭菌黑泥和辐照灭菌泥样品中分别检出8个和3个古菌门。进一步分析比较门水平下的物种相对丰度可知(图4),2种黑泥样品的优势门组成差别明显。BM0中平均相对丰度值较高的前2个门占古菌总测序数的99%,分别为1种未知古菌门(unclassified_k_norank_d__Archaea,57.53%)和盐杆菌门(Halobacterota,42.14%)。BM-CIS中平均相对丰度值最高的为嗜盐菌门(Halobacterot,88.45%),其次是热原体门(Thermoplasmatota,6.18%)和一种未知菌门(unclassified_k__norank_d__Archaea,5.37%)。Lusa等[27]研究发现,芬兰西海岸沼泽中的古菌群落属于泉古菌门(Crenarchaeota)、盐杆菌门和热原体门。Megyes等[28]利用宏基因组学方法研究罗马尼亚Beciu火山泥的细菌和古菌多样性,结果显示,该火山泥样的古菌群落以盐杆菌门和纳米古菌门(Nanoarchaeota)为主。
2.4.2 属水平组成
在属水平上,2种黑泥样品中分别检出50个和22个古菌属。对其属水平下的物种相对丰度进一步分析比较可知(图5),2种黑泥样品的优势属组成差别明显。BM0中平均相对丰度值较高的前5个属分别为1种未知古菌属(unclassified_k__norank_d__Archaea,57.53%)、其他属(others,22.09%)、Halovivax(7.02%)、Halorubellus(4.29%)和Haloterrigena(3.80%)。BM-CIS中各优势古菌属均匀度较高,平均相对丰度值较高的前5个属分别为其他属(others,28.53%)、Halomarina(13.80%)、unclassified_o__Halobacterales(9.64%)、norank_f__Halobacteriaceae(7.94%)和Haloferax(7.54%),与BM0均不相同。
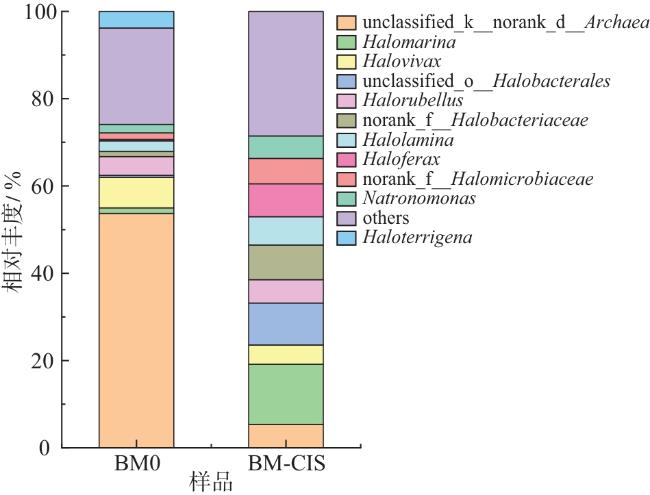
在属水平上,发现多种嗜盐细菌,如Halovivax、Haloterrigena、Natronomonas、Halanaeroarchaeum、Halorubellus、Halolamina和Halorussus,其在2种黑泥中的相对丰度及相关文献报道如表3所示。经过Co-60辐照灭菌处理后,黑泥中的一些嗜盐古菌被灭除,而一部分优势嗜盐古菌被保留。除了Halorussus之外,其余6种嗜盐古菌均为首次在山西运城盐湖中发现。
表3 在属水平上发现的多种嗜盐古菌 |
| 嗜盐古菌属名 | 相对丰度/% | 文献报道地区 | 文献 | |
|---|---|---|---|---|
| BM0 | BM-CIS | |||
| Halovivax | 7.02 | 4.38 | 伊朗Aran-Bidgol盐湖,中国内蒙古锡林浩特盐湖 | [29-30] |
| Halorubellus | 4.29 | 5.34 | 中国威海港溪海洋太阳盐沼,柴达木盆地可可盐湖 | [31-32] |
| Haloterrigena | 3.80 | - | 中国云南盐矿,美国加利福尼亚州太阳盐 | [33-34] |
| Halolamina | 2.43 | 6.50 | 沙特阿拉伯吉达南部海岸,墨西哥索诺拉盐田 | [35-36] |
| Halanaeroarchaeum | 2.03 | - | 俄罗斯阿尔泰库伦达盐湖 | [37] |
| Natronomonas | 1.93 | - | 韩国贡草湾,中国青海盐湖 | [38-39] |
| Halorussus | 2.14 | - | 山西运城盐湖 | [40] |
|
2.5 黑泥微生物群落结构的PCA分析
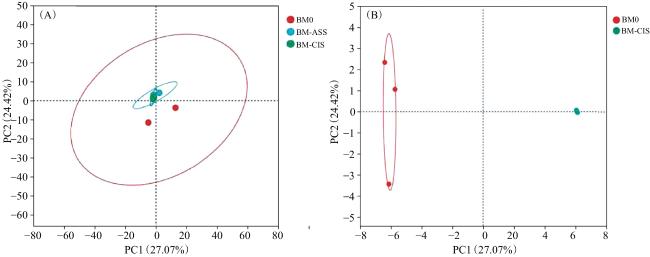
为分析不同灭菌方式对黑泥微生物群落结构的影响,对3种黑泥样品细菌和古菌属水平的群落结构进行PCA分析,样本微生物群落组成越相似,PCA图中的距离越近[24]。由图6A可知,BM0与BM-ASS、BM-CIS样本点距离较近,说明未灭菌黑泥与经灭菌处理后的2种黑泥所含细菌群落结构相似性较高。其中,BM-ASS和BM-CIS的聚类都包含在BM0中,且范围大幅减小,表明无论是高压蒸汽灭菌法还是Co-60辐照灭菌法的灭菌效果均较好;同时,BM-CIS的聚类包含在BM-ASS中,表明Co-60辐照灭菌法对细菌的总体灭除程度强于高压蒸汽灭菌法。由图6B可知,BM0和BM-CIS所含古菌群落产生明显的分离效应,表明这2种黑泥古菌属水平群落结构差异较大。BM0的3个样本点距离较远,分布离散,说明组内群落结构差异较大;而BM-CIS的3个样本点距离较近,且有重叠现象,说明组内群落结构差异较小。再次证明了不同灭菌方式对黑泥细菌和古菌菌群结构的影响不同,2种灭菌方式均可灭除大部分细菌,且Co-60辐照法对细菌的灭除程度强于高压蒸汽法;高压蒸汽法可以完全灭除古菌,而Co-60辐照法则可以灭除大部分古菌,保留部分嗜盐古菌。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
微生物群落结构的PCA分析结果表明,BM0样本点分布离散,造成其组内群落结构差异较大的原因可能是平行样本之间的盐度不同。郭浩然等[4]对运城盐湖6个采样点的18个土壤样品进行细菌多样性及影响因素分析,结果表明,因各盐湖理化成分差异,造成各土壤微生物的优势类群比例有所不同。本研究利用16S rRNA高通量测序方法对山西运城盐湖黑泥中的微生物多样性和群落结构,以及高压蒸汽灭菌和Co-60辐照灭菌2种灭菌方式对其多样性和结构的影响进行研究。从BM0中共检测出细菌29门76纲363属,古菌8门9纲50属;细菌优势菌群为变形菌门、无色杆菌属,古菌优势菌群为一种未知古菌门、一种未知古菌属;进一步分析,在细菌属水平上发现2种嗜盐细菌,其中化能自养硫氧化细菌(Guyparkeria)首次于运城盐湖中发现;在古菌属水平上发现多种嗜盐菌,其中Halovivax、Halorubellus、Halolamina、Halanaeroarchaeum、Natronomonas和 Haloterrigena首次在运城盐湖中发现。经过灭菌处理后,黑泥细菌和古菌群落丰富度和多样性均大幅度减少,但不同灭菌方式对黑泥微生物菌群结构的影响不同。对于细菌,2种灭菌方式均可以灭除大部分,且Co-60辐照法对细菌的灭除程度强于高压蒸汽法;对于古菌,高压蒸汽法可以完全灭除,而Co-60辐照法可以灭除大部分,保留部分嗜盐古菌。本研究为运城盐湖黑泥微生物的后续开发与利用提供参考。后期可结合黑泥的理化指标分析不同样品间微生物群落多样性。