

化学感受系统在昆虫觅食、求偶、产卵以及躲避天敌和有毒物质等生物学过程中起着关键作用[1-3]。昆虫主要借助触角等嗅觉器官对寄主植物进行远距离定位,依靠口器、足等化学感受器官进行近距离食物评估[4-5]。大量研究证实,部分鳞翅目昆虫降落于植物表面后,常以足碰触寄主叶片,感知其表面的信息化学物质,进而评估食物的安全性和可食用性,甚至判断该寄主植物是否适宜其幼虫取食[6-8]。因此,昆虫的足在味觉感知、寄主植物选择和产卵偏好中发挥着重要作用。
昆虫足化学感受功能的实现主要依赖于几个功能上相互关联的基因家族,包括气味结合蛋白(Odorant binding proteins,OBPs)、化学感受蛋白(Chemosensory proteins,CSPs)、气味受体(Olfactory receptors,ORs)、味觉受体(Gustatory receptors,GRs)、离子型受体(Ionotropic receptors,IRs)和感觉神经元膜蛋白(Sensory neuron membrane proteins,SNMPs)等[9-11]。杨叶青等[12]研究发现,中红侧沟茧蜂(Microplitis mediator)OBP19主要在前足跗节的感器下方表达,能与非挥发物和挥发物结合,参与味觉识别过程。Hua等[13]研究表明,在绿盲蝽(Apolygus lucorum)雄虫足中高表达的气味结合蛋白AlucOBP9与植物挥发物有较强的结合能力,暗示绿盲蝽足部参与了近距离识别化学信息物质的过程。果蝇足中的多种化学感受相关蛋白与求偶行为相关,如在果蝇雄虫前足特异表达的GR68 a直接参与雌虫性信息素的识别[14]。可见,这些基因家族在化学感受过程中发挥着不同作用,深入研究昆虫化学感受相关蛋白对揭示昆虫化学感受机理具有重要作用。
小菜蛾(Plutella xylostella)是十字花科植物专食性的农业害虫。作为鳞翅目昆虫,其雌雄成虫利用其灵敏的化学感受系统完成各自重要的生命活动[15-16]。小菜蛾雌虫将异硫氰酸酯类物质作为嗅觉信号,用于远距离寻找寄主植物,当其降落到寄主植物所在位置后,通过触角或足触碰叶片,以芥子油苷作为味觉信号刺激自身产卵[17-19];而小菜蛾雄虫则以雌虫释放的性信息素为信号,准确定位雌虫并与之交配,进而维持种群繁衍[20]。为初步解析小菜蛾雌、雄成虫足在化学感受过程中的分子机制,本研究收集了小菜蛾雌、雄成虫足样品,利用Illumina NovaSeq 6000测序系统进行转录组测序,将获得的序列进行拼接和组装,并进行基因功能注释和功能分类,深入挖掘其中的差异表达基因,获得了与小菜蛾化学感受功能相关的差异表达基因,为进一步了解小菜蛾感知外界信息化学物质的机制提供参考。
1 材料与方法
1.1 试验材料
供试昆虫为小菜蛾,其幼虫和成虫分别以新鲜甘蓝叶片和15%蜂蜜水饲养。饲养条件:温度(25±1)℃、相对湿度60%~75%、光周期14 L:10 D。
1.2 试验方法
1.2.1 总RNA的提取及质量检测
分别选取100只活动能力强的小菜蛾雌、雄成虫,置于解剖镜下,剪取足,液氮速冻后置于-80 ℃冰箱中备用。采用Trizol法提取上述样品的总RNA,经琼脂糖凝胶电泳和Agilent 5400检测RNA纯度,NanoDrop分光光度计检测RNA浓度,Agilent 5400检测RNA完整性,待RNA合格后由北京诺禾致源科技有限公司进行转录组文库构建和测序。
1.2.2 cDNA文库构建与测序
质检合格的总RNA首先通过Oligo(dT)磁珠富集带有poly A尾的mRNA,然后将其随机打断。以片段化的mRNA为模板,随机寡核苷酸为引物,在M-MuLV逆转录酶的作用下合成cDNA第一条链,然后用RNase H降解RNA链,在DNA polymerase I的作用下合成cDNA第二条链。纯化后的双链cDNA依次进行末端修复、加碱基A以及加测序接头操作,随后使用AMPure XP beads筛选长度在370~420 bp的cDNA进行PCR扩增,扩增产物经纯化后最终获得cDNA文库。文库构建完成后,先使用Qubit 2.0 Fluorometer进行初步定量,稀释文库至1.5 ng/μL,随后使用Agilent 2100对文库的插入片段大小进行检测,最后通过qRT-PCR对文库有效浓度进行准确定量,以保证文库质量。库检合格后,用Illumina NovaSeq 6000测序系统进行高通量测序。
1.2.3 转录组拼接和功能基因注释
为保证数据分析的质量及可靠性,转录组原始数据去除接头序列、poly N(N表示无法确定碱基信息)序列和低质量序列,计算Q20(clean data质量值≥20的碱基所占比例),Q30(clean data质量值≥30的碱基所占比例)和GC含量(clean data中G与C这2种碱基占4种碱基的百分比)。利用Trinity软件对高质量的clean data进行序列拼接与组装。组装得到的功能基因(unigenes)在NCBI非冗余蛋白数据库(Non-redundant protein sequences,NR)、核酸序列数据库(Nucleotide sequences,NT)、蛋白家族数据库(Protein family database,PFAM)、蛋白序列数据库SwissProt、GO数据库(Gene Ontology)、KOG数据库(真核生物蛋白质直系同源簇,Clusters of orthologous groups for eukaryotic complete genomes)和KEGG数据库(Kyoto encyclopedia of genes and genomes)7个数据库中进行BLAST比对,获得基因的功能注释信息。
1.2.4 基因差异表达分析
利用DESeq2软件计算转录组间的基因差异表达量,并进行显著差异性检验。将|log2FC|≥1且P≤0.05设定为显著差异表达的阈值,从中筛选出小菜蛾雌、雄成虫足间的差异表达基因。通过FPKM法对样品中的基因表达水平进行分析。
1.2.5 差异基因表达的GO和KEGG分析
利用GOseq 1.10.0软件对小菜蛾雌、雄成虫足间的差异表达基因进行GO功能富集分析,利用KOBAS v2.0.12软件对差异表达基因进行代谢通路注释。
1.2.6 关键基因的鉴定
为明确小菜蛾雌、雄成虫足间差异表达基因中是否存在化学感受基因,在筛选出的差异表达基因中,通过使用不同化学感受基因家族的基因全称和缩写以及通过BLAST比对进行查找,并在转录组数据库中查找编码这些基因的序列,以确定关键基因。
2 结果与分析
2.1 转录组序列分析和组装
前期研究发现,小菜蛾雌、雄成虫足转录组数据过滤后碱基数均≥6 GB,GC含量均>45%,Q20比例均>96%,Q30比例均>90%,说明此次测序结果可靠,可用于后续的拼接组装[21]。
由表1可知,原始序列经过拼接与组装,共得到141 226个转录本,其中长度在1 kb以上的转录本有54 281个,序列平均长度为1 175 bp,N50长度为1 707 bp。转录本进一步组装获得46 554个unigenes,总长度为51 410 982 bp,平均长度为1 104 bp,N50长度为1 791 bp。
表1 小菜蛾成虫足测序数据的组装质量结果 |
| 项目 | 转录本 | 功能基因 | |
|---|---|---|---|
| 不同长度范围 的数量/bp | <500 | 42 965 | 18 195 |
| 500~1 000 | 43 980 | 13 648 | |
| 1 000~2 000 | 32 981 | 7 989 | |
| >2 000 | 21 300 | 6 722 | |
| 总数/个 | 141 226 | 46 554 | |
| 平均长度/bp | 1 175 | 1 104 | |
| N50值/bp | 1 707 | 1 791 | |
| 总长度/bp | 165 903 141 | 51 410 982 | |
2.2 基因功能注释
由表2可知,所有unigenes与NR、NT、KEGG、SwissProt、PFAM、GO和KOG 7个数据库进行比对,共有46 554条unigenes成功注释。其中,NT数据库中注释到的unigenes最多(31 074条,占比66.75%),KOG数据库中注释的最少(8 552条,占比18.37%)。
表2 小菜蛾功能注释结果统计 |
| 数据库 | unigenes数量/条 | 注释比例/% |
|---|---|---|
| NR数据库注释 | 24 879 | 53.44 |
| NT数据库注释 | 31 074 | 66.74 |
| KEGG数据库注释 | 11 737 | 25.21 |
| SwissProt数据库注释 | 15 564 | 33.43 |
| PFAM数据库注释 | 16 191 | 34.77 |
| GO数据库注释 | 16 189 | 34.77 |
| KOG数据库注释 | 8 552 | 18.37 |
| 所有数据库均有注释 | 5 009 | 10.75 |
| | 35 314 | 75.85 |
| 总基因数 | 46 554 | 100 |
2.3 基因功能分类
前期研究发现,小菜蛾雌、雄成虫足转录组共有16 189条unigenes注释到GO数据库中,注释到生物过程的unigenes有33 945条,其中细胞过程(9 122条)和代谢过程(7 781条)数量较多;注释到细胞组分的unigenes有14 974条,其中细胞结构体(7 041条)和细胞内组分(4 043条)数量较多;注释到分子功能的unigenes有18 609条,其中unigenes数量较多的为结合活性(8 749条)和催化活性(6 233条)[21]。
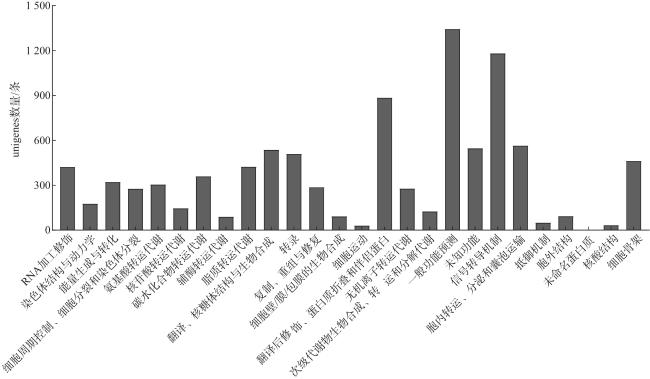
由图1可知,小菜蛾雌、雄成虫足转录组共有8 552条unigenes注释到26个KOG分类中。其中一般功能预测的unigenes数量最多,为1 345条;其次为信号转导机制,为1 182条;未命名蛋白数量最少,仅有1条。
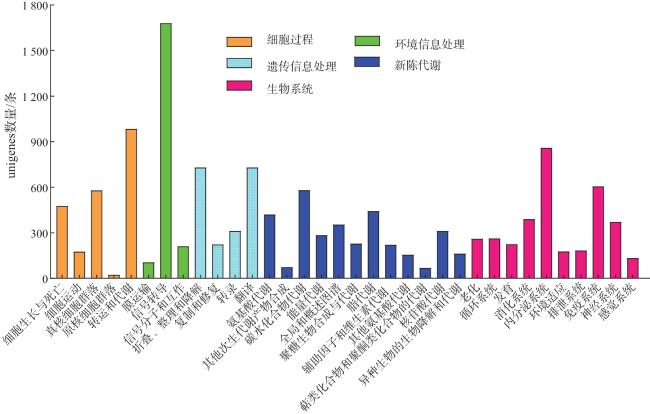
由图2可知,KEGG数据库的注释结果显示,11 737条unigenes注释到5个通路34个二级分支中,其中环境信息处理通路中参与信号转导相关的unigenes数量最多,为1 682条。
2.4 差异表达基因的筛选
2.5 差异表达基因GO富集分析
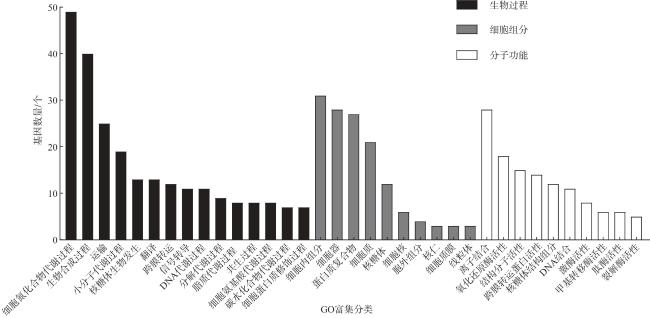
共118个差异表达基因注释到GO数据库中,并被富集到生物过程、细胞组分和分子功能3大类88个亚类中,分别包含16、42和30个亚类。由图4可知,在生物过程中,差异表达基因在细胞氮化合物代谢过程(49个)所占比例最高;在细胞组分中,差异表达基因在细胞内组分(31个)、细胞器(28个)和蛋白质复合物(27个)这3个亚类中所占比例较高;分子功能中,差异表达基因在离子结合(28个)中的占比最高。
2.6 差异表达基因KEGG代谢通路分析
KEGG代谢通路分析发现在255个差异表达基因中,有74个差异表达基因参与125条不同的代谢通路,其中62个基因上调表达,12个基因下调表达。由表3可知,差异表达基因主要富集在核糖体(16个)、寿命调节通路(12个)、酪氨酸代谢(5个)、角质素、软木脂和蜡质生物合成(4个)、泛醌等萜醌类生物合成(3个)和苯丙氨酸代谢(3个)。
表3 差异基因KEGG富集通路(前10个) |
| 序号 | KEGG分类 | 基因数量/个 |
|---|---|---|
| 1 | 核糖体 | 16 |
| 2 | 寿命调节通路 | 12 |
| 3 | 酪氨酸代谢 | 5 |
| 4 | 角质素、软木脂和蜡质生物合成 | 4 |
| 5 | 泛醌等萜醌类生物合成 | 3 |
| 6 | 苯丙氨酸代谢 | 3 |
| 7 | 抗生素的生物合成 | 9 |
| 8 | 剪接体 | 6 |
| 9 | 苯乙烯降解 | 2 |
| 10 | 非酒精性脂肪性肝病 | 5 |
2.7 关键化学感受基因的鉴定
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
随着高通量测序技术的发展,转录组测序技术已成为一种重要的研究手段。转录组测序技术为物种或组织中所有转录产物的集合,能够反映不同物种、同一物种的不同组织以及同一组织不同时间基因的表达模式,有利于揭示昆虫形态、生理、行为以及特定生物学过程中的分子机理[22]。足是昆虫最重要的化学感受器官之一,目前其感受外界信息化学物质的分子机制尚未完全明确。本研究通过对小菜蛾雌、雄虫足进行转录组测序,共获得46 554条unigenes序列,平均长度1 104 bp,N50长度为1 791 bp,Q30比例大于90%。为了保证转录组测序的质量,Q30碱基比例应不低于80%,N50值越大表明序列拼接的完整性越好[23-25]。此外,本研究中基因功能注释显示,46 554条unigenes均被成功注释。由此可见,试验所获得的转录组数据质量良好,保证了后续分析的准确性和全面性。
对基因的功能进行分类有助于系统地认识基因的功能,掌握基因的产物及其在生命活动中的作用。小菜蛾足转录组有16 189条unigenes富集到GO数据库中,分别隶属于生物过程、细胞组分与分子功能三大类,其中注释到生物过程的unigenes数量最多,其次为分子功能。在生物过程中,参与细胞过程和代谢过程的unigenes数量最多,说明小菜蛾足中的细胞新陈代谢活跃。在分子功能中,具有结合活性和催化活性的unigenes数量最多,说明足中的许多基因产物和蛋白质具有结合和催化作用。注释到KOG数据库中的unigenes根据功能被分为26类,其中注释到只有一般功能预测的unigenes数量最多,这与许多昆虫的转录组研究结果一致[26-28]。与此同时,小菜蛾足转录组数据经KEGG注释的代谢通路中,参与信号转导的unigenes数量最多。以上数据充分说明小菜蛾足中的许多unigenes可启动或激活相关的细胞信号转导途径影响细胞通信,进而使昆虫表现出整体的生物学效应[29]。
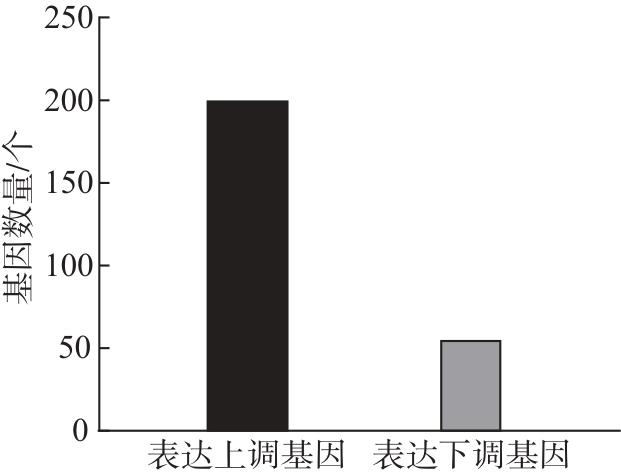
足中的化学感受系统是昆虫感知、处理与整合外界信息化学物质的重要信号转导途径之一。为明确小菜蛾雌、雄足在化学感受过程中的分子机制,差异表达基因筛选结果显示,小菜蛾足的255个基因在雌、雄虫中差异表达,其中200个基因表达水平显著上调,55个基因表达水平显著下调。GO富集分析发现,这些差异表达基因在分子功能方面主要富集在离子结合,说明这些基因产物具有结合功能。KEGG代谢通路分析有74个差异表达基因参与125条不同的代谢通路,这些通路主要是调节、生物合成和物质代谢的通路。
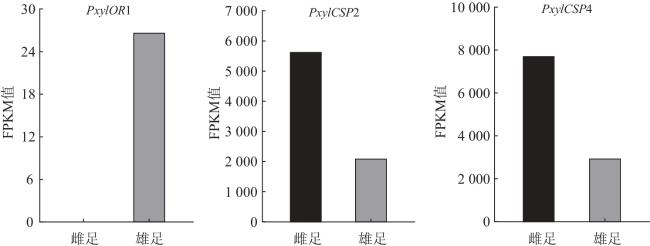
PxylCSP2和PxylCSP4在小菜蛾触角、头、腹、足和翅中均有表达,其中PxylCSP2在触角和头中的表达量最高,PxylCSP4在各组织中表达量差异不显著[32]。本研究中,PxylCSP2和PxylCSP4在小菜蛾雌虫足中的表达量高于雄虫足。CSPs是昆虫化学感受通路的重要组成部分,被认为是昆虫感知外界化学信息物质的第一步,其功能具有多样性,其最基本的功能为结合并运输化学信息物质,如寄主植物挥发物、性信息素和报警信息素等[33-36]。此外,相关研究表明,在美洲大蠊(Periplaneta americana)触角和头中表达的PamerCSP在足再生的过程中表达量显著增加,表明其与组织再生相关[36-37]。因此,PxylCSP2和PxylCSP4具有化学感受功能还是组织再生功能值得进一步研究。
综上,本研究在小菜蛾雌、雄成虫足转录组中共获得46 554条unigenes序列,筛选得到255个差异表达基因,其中3个化学感受基因(PxylOR1、PxylCSP2和PxylCSP4)在雌雄足中差异表达,为小菜蛾足中功能基因的研究提供重要信息,为揭示昆虫感知外界信息化学物质的机制提供理论依据。