

茄子(Solanum melongena L.)是茄科茄属作物,其果实可作为蔬菜食用,根、茎、叶可入药,具有较高的经济价值。青枯病是茄子主要的土传病害之一,致使茄子减产20%~30%,严重时损失在50%~60%[1],一定程度上制约茄子产业的可持续健康发展。茄子青枯病的病原菌为青枯劳尔氏菌(Ralstonia solanacearum),又称青枯假单胞菌,属薄壁菌门劳尔氏菌属,是一种细菌性病害。病原在寄主体内可存活半年,在土壤中则可存活14个月至6年之久。病原主要在土壤中越冬,第2年随雨水、灌溉水及土壤传播,从寄主根部或茎基部伤口侵入。发病初期仅个别枝上部的几片叶片颜色变淡,呈现局部萎垂,随后下部叶片凋萎,中部叶片凋萎最迟,后期因叶绿素逐渐分解而变为黄色或褐色,最后焦枯。该病害始于茎基部,剖开病茎基部发现木质部变褐色,以后延伸至枝条。感染青枯病的茄子茎段和枝条的髓部大多溃烂或中空,湿度大时,用手挤压病茎横切面,会有少量乳白色黏液溢出。该病害易在高温高湿、重茬连作、田间积水和偏施氮肥等情况下发生。郑雪芳等[2]研究表明,茄子青枯病发病率与气温、地温、相对湿度、降水量、光照辐射和二氧化碳浓度呈极显著正相关。青枯病的菌群分布特性为病株及其根际土壤分布数量显著多于健康植株及其根际土壤,分布数量为土壤>根>茎[3]。蓝江林等[4]研究发现,茄子青枯病菌在根部分离到的内生细菌最多,其次为茎部,叶部最少。
1 材料与方法
1.1 试验地基本情况
试验地位于浙南一茄果类种苗繁育基地,已连续繁育茄果类种苗5年,该基地属亚热带季风性湿润气候,四季分明,温暖湿润,年日照时数1 785.7~3 118.6 h,年太阳辐射总量5 040~5 400 MJ/m2,年平均气温17.0 ℃,无霜期259 d,年降水量1 537 mm。
1.2 试验材料
试验材料为茄子品种紫红04,育苗基质来自农资公司,成分为泥炭、椰糠、有机肥和珍珠岩等,所用基质经北京芸归生物科技有限公司权威检测,发现根际基质中含有青枯劳尔氏菌。
1.3 试验设计
7月开展茄子育苗工作,经20 d育苗,发现茄子苗大面积感病,主要症状表现为轻病株叶片卷曲、萎蔫,重病株茎秆出现水渍直至全株枯萎腐烂,发病植株茎秆横切面用手挤压可见乳白色液体,纵切面维管束变褐色,与青枯病症状相似。设置感病的茄子植株为Ts处理,取对照育苗基地(育苗基质中不含有机肥)生长正常的茄子植株为CKs处理,各处理设置3个重复,完成植株茎秆内生细菌群落结构及多样性检测与分析。
1.4 样品采集与处理
在育苗20 d后,各处理分别选取3棵感病和未感病的茄子植株,取植株茎秆3 cm长度用于检测,其中Ts处理茎秆截取病健交界处组织,之后采用表面消毒研磨法进行样品处理。
1.5 测定方法
1.5.1 植株茎秆微生物基因组DNA的提取及检测
首先对研磨后的茎秆样品采用天根试剂盒(DP712)提取总DNA,之后使用Nanodrop检测DNA样品浓度及纯度;琼脂糖凝胶电泳检测DNA样品完整性。取适量样本DNA于离心管中,使用无菌水稀释样本至1 ng/μL。使用前端引物:341F-5’-CCTACGGGNGGCWGCAG-3’;后端引物:806R-5’-GACTACHVGGGTATCTAATCC -3’对目标区域(V3~V4)进行PCR扩增。以稀释后的基因组DNA为模板,根据测序区域的选择,使用带Barcode的特异引物、Buffer和高效高保真酶进行PCR,确保扩增效率和准确性。
1.5.2 PCR产物的混样和纯化
PCR产物使用2%浓度的琼脂糖凝胶进行电泳检测;根据PCR产物浓度进行等量混样,充分混匀后使用2%琼脂糖凝胶电泳纯化PCR产物,剪切回收目标条带。
1.5.3 文库构建和上机测序
使用建库试剂盒进行文库的构建,将构建好的文库经过Qubit定量和文库检测合格后,采用16S rRNA基因高通量测序技术,通过Illumina NovaSeq平台进行上机测序,所测数据经过去除嵌合体序列处理后得到有效序列。
1.6 数据分析
测序后对原始数据进行拼接、过滤、去除嵌合体处理,得到优化序列,进行Alpha多样性分析、物种组成分析及物种差异分析。用Chao1指数(物种丰富度)和Shannon指数(物种多样性)两项指标指示多样性。试验数据均采用Microsoft Excel 2010软件进行统计分析。
2 结果与分析
2.1 茄子茎秆内生细菌Alpha多样性分析
稀释曲线用于验证测序数据量能够反映样品中的物种多样性,并间接反映样品中物种的丰富程度。持续抽样下新物种出现的速率见图1,在0~150 000条范围内,随着测序条数的加大,曲线趋于平缓,表示此环境中的物种并不会随测序数量的增加而明显增多,说明此次测序可反映感病和未感病植株茎秆细菌群落结构多样性的真实情况。
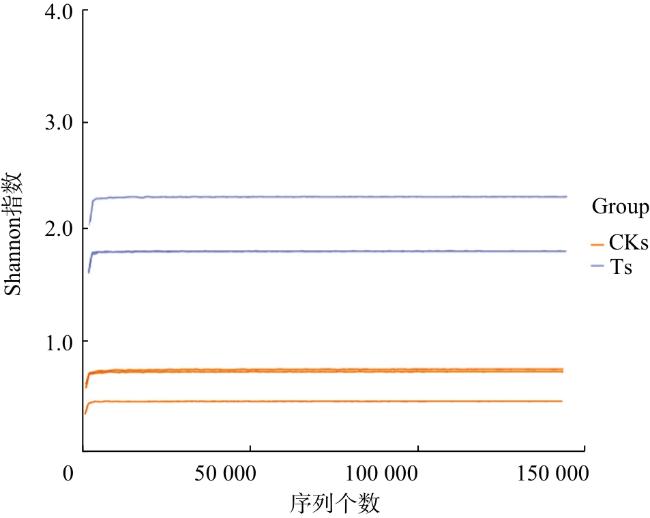
由表1可知,样品有效序列总数在188 979~210 091个。在97%相似度水平下,样品细菌OTUs总数在179~454,覆盖度均在99.0%,说明测序文库已经达到了饱和状态。
表1 感病与未感病茄子茎秆内生细菌Alpha多样性特征 |
| 处理 | 多样性指数 | ||||
|---|---|---|---|---|---|
| 有效序 列/条 | OTUs总数 | Chao1指数 | Shannon指数 | 测序深度 | |
| CKs1 | 194 117 | 454 | 681 | 0.75 | 0.99 |
| CKs2 | 199 888 | 179 | 262 | 0.41 | 0.99 |
| CKs3 | 210 091 | 339 | 528 | 0.70 | 0.99 |
| Ts1 | 204 724 | 188 | 269 | 1.81 | 0.99 |
| Ts2 | 209 145 | 253 | 318 | 2.33 | 0.99 |
| Ts3 | 188 979 | 390 | 519 | 1.82 | 0.99 |
|
由表1可知,CKs和Ts的平均Chao1指数分别为490.33、368.67,Ts的平均Chao1指数比CKs低24.81%,说明感病植株内生细菌的物种丰富度低于健康植株。从Shannon指数可知,Ts处理比CKs高220.43%,说明感病植株内生细菌的物种多样性高于健康植株。由此表明,感病植株相比健康植株降低了细菌菌群丰富度,提高了细菌菌落多样性。
2.2 茄子茎秆内生细菌门水平分类组成
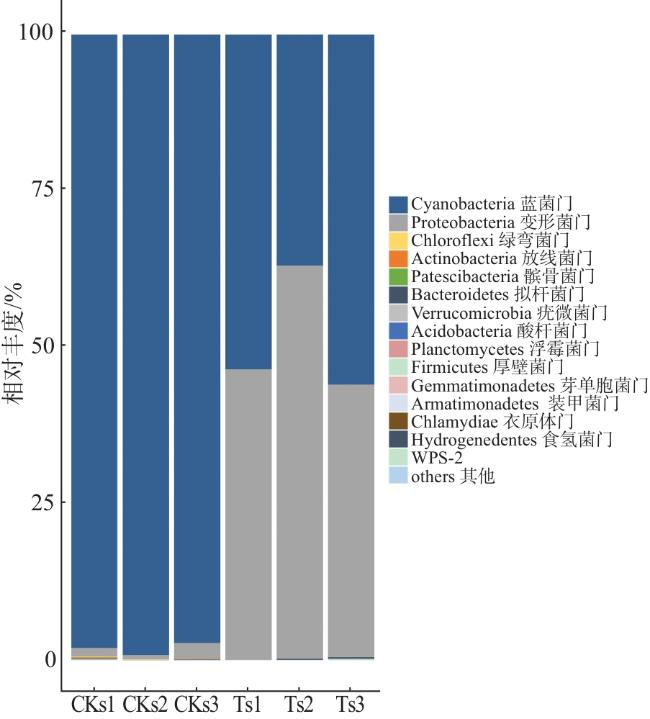
由图2可知,感病与未感病茄子茎秆内生细菌排名前3的优势门类分别是蓝菌门(Cyanobacteria)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)。其中,未感病茄子茎秆中蓝菌门的相对丰度(98.16%)较感病茎秆(49.05%)高49.11个百分点;感病茄子茎秆中变形菌门的相对丰度(50.68%)较未感病茎秆(1.49%)高49.19个百分点;感病与未感病茄子茎秆中绿弯菌门的相对丰度分别为0.006%、0.080%。
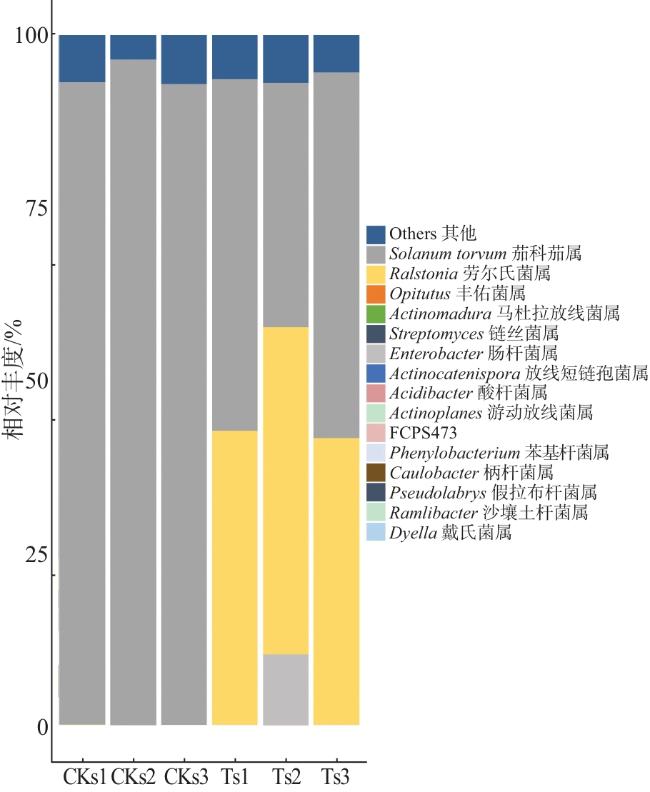
2.3 茄子茎秆内生细菌属水平分类组成
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
生物多样性是衡量生态系统稳定和健康的一个重要指标,适宜的生长环境表现为多样性和均匀度高、优势度低[8]。其中,植株内生菌多样性是植物内生菌与植物间微生态关系平衡的重要保障[9]。植物内生细菌的群落结构与多样性不仅与植物种类、基因型有关,还与植物的生长阶段、感病情况及环境因素等有关[4]。Reiter等[10]研究发现,马铃薯的内生细菌群落结构与病原菌的发生密切相关,被胡萝卜软腐欧文氏菌马铃薯黑胫亚种(Erwiniacarotovora subsp. atroseptica)侵染的马铃薯病株内生细菌群落多样性高于健康植株。孙美丽等[11]研究认为,感病的烟叶细菌多样性指数高于健康烟叶,而丰富度指数低于健康烟叶。本研究结果与其较一致,本研究发现,感病植株相比健康植株,内生细菌菌群丰富度明显降低,菌落多样性明显提高,表明感病植株茎秆内生细菌菌群趋于饱和。而蓝江林等[4]研究认为,感病植株的内生细菌的群落多样性要低于健康植株,可能是因为取样的状态不同,本研究采集的是中度感染青枯病的病株样本,而蓝江林等[4]采集的病株已完全表现为青枯病症状,病株体内的内生细菌已占据数量上的绝对优势,导致病株内生细菌多样性下降。
植物内生微生物是指能够在健康植物器官或组织中生存而不引起寄主植物病变的一大类微生物,主要包括细菌、真菌和放线菌等[12-13]。其中,内生细菌是植物内生微生物的重要组成部分,具有促生、增强宿主抗性及生物防治等功效。微生物群落结构是指在特定环境中定植的各种微生物组成的一个生态系统,其会受到环境因子的影响,不同微生物菌群也会受一个或多个环境因子的调控[14]。蓝菌门也被称为蓝绿藻,分布于海水、土壤和树干等载体中,其中一部分分布于植物体内,形成内生植物,是一类能透过产氧光合作用获取能量的细菌。蓝细菌含有叶绿素a和b,是一种能够在光合作用下释放氧气的原核微生物,主要以二分裂或多分裂方式进行繁殖,少数蓝细菌可形成孢子,抵抗不良环境。本研究发现,未感病茄子茎秆蓝菌门的相对丰度为98.16%,感病茎秆只有49.05%。说明感病茎秆内生细菌蓝菌门的相对丰度比未感病的茎秆低。变形菌门是一类广泛存在于自然界中的细菌,包括一些致病菌和有益菌。变形菌门包括多种病原菌,如大肠杆菌、沙门氏菌、霍乱弧菌、幽门螺杆菌等,也有自由生活的种类,包括很多可以进行固氮的细菌,参与土壤有机物分解,进行光合作用等。本研究发现,感病茄子茎秆变形菌门的相对丰度为50.68%,未感病茎秆变形菌门的相对丰度为1.49%,感病茄子茎秆变形菌门的相对丰度比未感病茎秆高49.19个百分点。可能是由于茄子感染青枯病,而病原菌青枯雷尔氏菌属于细菌界,变形菌门、变形菌纲(Betaprotebacteria)、薄壁菌目(Graeilieutes)、假单胞菌科(Pseudomonaeeae)、劳尔氏菌属(Ralstonia),是极具暴发性和破坏性的细菌病原体之一[15-17]。因此,感病茄子茎秆变形菌门的相对丰度远高于未感病植株茎秆。
本研究发现,未感病茄子茎秆茄科茄属的相对丰度为94.12%,比感病茎秆高47.68个百分点,说明感病茄子茎秆茄科茄属的相对丰度受到其他因素的抑制作用。茄科劳尔氏菌是一种危害十分严重的植物病原细菌,其引起的植物青枯病严重影响番茄、茄子、辣椒等作物的健康生产。该病菌可同病株残体一同进入土壤,长期生存形成侵染源,病原菌在土壤中并非以休眠状态生存,而是在发病植株或杂草的根际进行繁殖。病原菌主要是由作业过程中造成的伤口或幼虫等根部害虫造成的伤口侵染植株,在茎的导管部位和根部发病。本研究发现,感病茄子茎秆劳尔氏菌属的相对丰度为43.89%,未感病的茄子茎秆劳尔氏菌属的相对丰度为0.01%,二者相差43.88个百分点。由此可见,感病茎秆劳尔氏菌属的相对丰度占比接近50%,接近中度感染状态。
本研究为探讨茄子青枯病对植株茎秆内生细菌群落的影响提供了初步数据,但研究存在一个重要局限性:在属水平分析中,误将植物茄科茄属注释为细菌属并纳入统计,这很可能源于测序过程中16S rRNA同源序列的非特异性扩增,或所用参考数据库存在污染。这使得属水平的丰度比较缺乏可靠性,也可能干扰Alpha多样性指数的解读。尽管此问题影响了属水平分析的准确性,但门水平等的核心结果仍具有明确的生物学意义。未来研究需通过优化引物特异性或在生物信息学流程中严格过滤非细菌序列,以提高数据解读的可靠性。
综上,采用16S rRNA基因组高通量测序的方法,分析了感染青枯病和健康植株茎秆内生细菌群落的变化特征。结果表明,相比健康茄子植株,感染青枯病的植株茎秆降低了内生细菌菌群丰度和有益菌丰度,提高了菌落多样性和有害菌丰度。本文为后期茄子青枯病在基质或土壤修复上的预防提供参考。