

大黄鱼(Larimichthys crocea)是重要的海水经济鱼类,2024年养殖产量达28万t[1]。其肉质鲜嫩,富含蛋白质、脂肪、微量元素以及无机盐等,是理想的膳食来源之一[2]。随着人工养殖种质退化与养殖环境的变化,大黄鱼出现了肉质松软、风味下降等品质退化的特征,对水产养殖产业的可持续发展造成了不利影响[3]。鲜味是肉质风味的重要组成部分,除氨基酸外,核苷酸类物质也是养殖动物肉品鲜味的重要来源[4]。水产动物的主要鲜味核苷酸是肌苷酸(Inosine 5'-monophosphate,IMP),其鲜味强度是谷氨酸钠的40倍,是衡量肌肉品质的重要指标[5]。多种重要酶参与嘌呤核苷酸代谢,其中,腺苷酸激酶(Adenylate kinase,AK)家族同工酶参与细胞内核苷酸的合成,对肉品风味具有重要的调控作用[6]。
AK是一种单体磷酸转移酶,在二价化合物,如Mg2+的辅助下可逆催化腺苷三磷酸(ATP)与腺苷酸(AMP)生成腺苷二磷酸(ADP)[7]。AK通过上述反应影响细胞内核苷酸含量,从而调控三羧酸循环、氧化磷酸化和糖酵解等重要生命过程。AK具有多个亚型,其中,AK1在骨骼肌和脑组织的细胞质中表达[8]。Zhang等[6]研究发现,静原鸡肌肉组织中ak1基因的mRNA表达量与IMP含量呈正相关。目前,对于AK的研究,多集中于高等生物的细胞运动、疾病发生和细胞分化、凋亡等方面,关于其对水产动物IMP代谢的影响还有待进一步研究。本研究克隆了大黄鱼a k1基因(Lca k1)编码区(CDS)序列,解析了其序列与进化特征,并应用荧光定量PCR技术(qPCR)对该基因的组织表达进行分析,为探究AK1在核苷酸和能量代谢中的作用提供参考。
1 材料与方法
1.1 试验鱼类及前处理
试验大黄鱼取样自福建省宁德市三都澳海域养殖渔排,平均体长约21 cm,体质量约100 g。随机选取5尾大黄鱼,置于含MS-222(50 mg/L)的海水中麻醉30 s后,解剖并摘取脾脏、肝脏、头、肾、鳃、心脏、脑、肠、肌肉、皮肤、胃和肾脏组织,迅速转移至液氮中速冻,随后转移至超低温冰箱冻存,备用。
1.2 总RNA的提取与反转录
使用RNA提取试剂盒(Promega,USA)提取各组织的总RNA,经琼脂糖凝胶电泳检测总RNA的完整度后,应用Nano Drop微量核酸测定仪(Thermo,USA)测定总RNA的浓度。按照反转录试剂盒(Promega,USA)操作说明,配制反转录反应体系,获取各组织cDNA模板,-20 ℃冻存备用。
1.3 Lcak1基因CDS的克隆
根据NCBI预测的大黄鱼ak1基因序列(XM_019276271.2),应用Primer 5软件设计Lcak1基因CDS序列的PCR引物,Lcak1-F:5'-ATGGCAGACAAAATTAAAGATG-3',Lcak1-R:5'-CTACAGTGCATCAATAGCCGTAG-3'。以制备的肌肉组织cDNA为模板,PCR扩增目的基因,经浓度为2%的琼脂糖凝胶电泳检测后,使用DNA片段回收试剂盒(天根,中国)纯化和浓缩PCR产物后,按照pMD-19T(Takara,Japan)载体说明书配制连接体系,将目的基因经T-A克隆连接至载体中,将重组载体转化至大肠杆菌DH5α感受态细胞中并铺覆于含氨苄青霉素的固体LB培养基上,次日,使用载体通用引物经菌落PCR筛选阳性克隆后,送福州尚亚生物技术有限公司测序。
1.4 Lcak1基因的序列特性分析
对测序获得的Lcak1基因序列进行分析,使用ORF Finder工具(https://www.ncbi.nlm.nih.gov/orffinder/)分析Lcak1基因CDS序列并预测其氨基酸序列。使用蛋白质理化性质分析工具Expasy(https://web.expasy.org/compute_pi/)预测LcAK1蛋白的相对分子量大小与等电点;使用Signal P 5.0(https://services.healthtech.dtu.dk/services/SignalP-5.0/)和TMHMM 2.0(https://services.healthtech.dtu.dk/services/TMHMM-2.0/)分别预测LcAK1蛋白的信号肽和跨膜结构域;使用NetPhos 3.1(https://services.healthtech.dtu.dk/services/NetPhos-3.1/)分析LcAK1蛋白的磷酸化位点;使用NetNGlyc 1.0(https://services.healthtech.dtu.dk/services/NetNGlyc-1.0/)和NetOGlyc 4.0(https://services.healthtech.dtu.dk/services/NetOGlyc-4.0/)分别预测LcAK1蛋白的N-糖基化和O-糖基化位点。
1.5 LcAK1蛋白结构与进化分析
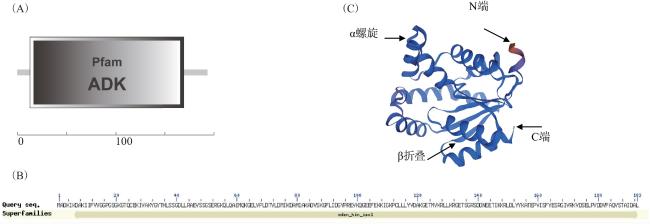
使用SMART(http://smart.embl-heidelberg.de/)和CD Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)在线工具分析LcAK1蛋白的结构域,使用SWISS-MODEL(https://swissmodel.expasy.org/)预测LcAK1蛋白的三维结构。对LcAK1进行BLAST(https://blast.ncbi.nlm.nih.gov/Blast.cgi)比对分析,选取不同种AK1的氨基酸序列,使用MEGA 7.0软件以N-J邻接法构建其系统进化树。
1.6 Lcak1基因表达模式分析
表1 Lcak1基因的qPCR引物 |
| 引物名称 | 引物序列(5'-3') | 基因编号 | 用途 |
|---|---|---|---|
| qLcak1-F | GAAGCGCCTGGACCTGTATT | XM_019276271.2 | 目的基因qPCR扩增 |
| qLcak1-R | CAATGCCTCGGCTCTCATAA | ||
| qLcβ-actin-F | GACCTCACAGACTACCTCATG | GU584189.1 | 内参基因qPCR扩增 |
| qLcβactin-R | TGTTGTAGGTGGTCTCGTGGA |
1.7 数据统计与分析
使用SPSS软件对Lcak1基因的mRNA在各组织中的相对表达量进行单因素方差(ANOVA)及Duncan多重比较分析,应用GraphPad Prism 8.0软件作图。
2 结果与分析
2.1 Lcak1基因CDS的克隆与序列特征
测序结果如图2所示,Lcak1基因的CDS片段长582 bp,编码193个氨基酸,对应LcAK1蛋白质分子量为21.24 kDa,等电点为6.62。经生物信息学软件分析,LcAK1不含信号肽,无跨膜结构域,共有15个磷酸化位点,包括9个丝氨酸(S)残基、5个苏氨酸(T)残基以及1个酪氨酸(Y)残基位点。此外,LcAK1蛋白无N-连接糖基化修饰位点,但在第135位氨基酸(T)含有1个O-糖基化修饰位点。

2.2 LcAK1蛋白结构与进化分析
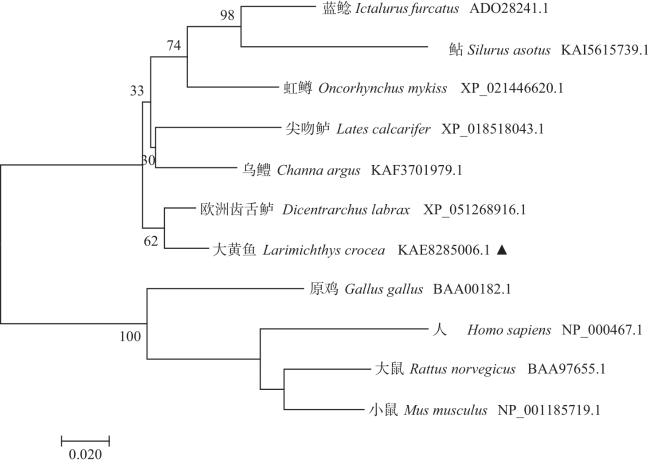
系统进化树显示,LcAk1与欧洲齿舌鲈(Dicentrarchus labrax)亲缘关系最近,与哺乳类动物最远。大黄鱼与欧洲齿舌鲈聚为一小支后,再与其他鱼类聚为一大支(图4)。
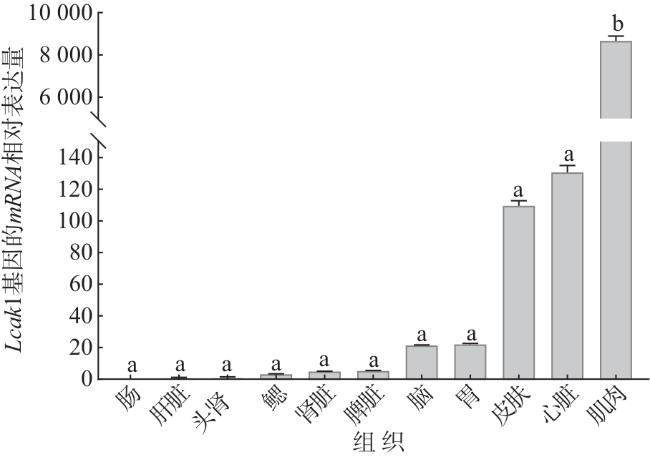
2.3 Lcak1基因的表达模式
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
核苷酸在生物体内具有多重生物学功能,既能作为高能化合物参与细胞能量代谢,也可作为活化中间体介导多种生物合成途径,还能作为信号分子参与细胞通信过程,其在维持细胞正常生理活动中发挥着不可或缺的作用[10]。AK通过磷酸基团的转移调节细胞内的不同腺嘌呤核苷酸比例,在维持细胞中核苷酸的含量和能量代谢活动中起着关键作用[11]。AK1是主要的细胞质亚型,其编码基因已在部分水产动物中被克隆。本研究克隆的大黄鱼ak1基因编码193个氨基酸,对应蛋白质分子量约21.24 kDa,这与牙鲆(Paralichthys olivaceus)[7]的ak1分子量大小相当。ak1基因在大部分组织中都有表达,但在骨骼肌、脑和红细胞中表达量最高,这可能与AK1广泛参与细胞能量代谢相关联[12]。本研究发现,Lcak1基因在肌肉组织中的表达量明显高于其他组织,这与高等生物一致;在心脏组织也有较高表达,推测与心脏组织中血液含量较高有关[13];此外,皮肤组织中也检测到较高的Lcak1表达,这可能与取样皮肤组织时未完全剔除肌肉有关,也可能与种属差异有关,具体原因有待进一步探究。
AK是机体嘌呤核苷酸代谢中的关键酶,其基因表达量与IMP含量相关。在鸡中发现,腿部肌肉组织中ak1基因的mRNA表达量与IMP含量呈正相关,表明AK1在IMP代谢中发挥关键调控作用,其对应基因可作为肉质风味改良的候选基因[6]。本研究仅对ak1基因在大黄鱼各组织中的分布进行了检测,未开展其基因表达量与肌肉IMP含量的关联性分析,后续拟对不同养殖规格大黄鱼进行取样,分别检测其肌肉IMP含量与ak1基因的表达量,以探究二者的相关性,明确ak1基因在调控大黄鱼肉质风味中的作用。