

土壤盐分是作物生长的主要环境制约因素之一,随着气候变化,部分水稻种植区的稻田盐碱度不断增加,影响了粮食生产。因此,提高水稻对盐碱胁迫的耐受性对保障粮食安全具有重要意义。硅是植物生长发育的非必需营养元素,近年来其在增强植物对非生物胁迫的耐受能力方面受到广泛关注。赵鑫等[1]研究指出,增施外源硅可通过降低电解质渗透率,增加番茄根、茎、叶中的铁含量,提高其对低铁胁迫的耐受能力。干旱胁迫下,施加外源硅可提高草坪草[2]、黄瓜[3]、小麦[4]的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性;降低小麦[5]、水稻[6]叶片的过氧化氢(H2O2)和丙二醛(MDA)含量,从而提高植物抗旱性。张倩等[7]研究表明,盐胁迫下棉花增施外源硅后,其耐盐性明显提高。闫国超等[8]研究指出,在100 mmol/L NaCl处理下,添加1.5 mmol/L硅,提高了水稻SOD、CAT、 抗坏血酸过氧化物酶(APX)的活性,降低了MDA含量,从而提高了水稻的耐盐性。本文通过测定水稻的生长指标和生理生化指标,分析外源硅增强水稻幼苗耐盐性的作用机制;并通过盆栽试验,对该调控效应进行室外实践验证。研究结果将为阐明硅缓解水稻盐害的生理生化机制及其实践应用提供参考,也为研发提高水稻耐盐能力的生态绿色优质肥料提供依据。
1 材料与方法
1.1 试验材料
利用引进水稻品种Nipponbare进行水培试验,用于室内耐盐差异性分析和各项生理生化指标的测定;利用连粳7号(苏审稻201008)和连粳15号(苏审稻20170006)进行田间盆栽试验验证。MDA含量测定试剂盒,以及CAT、SOD、APX和POD活力检测试剂盒均购自南京建成生物工程研究所。Na2SiO3、NaCl、浓硝酸试剂(优级纯)均购自国药集团化学试剂有限公司。试验仪器:酶标仪Multiskan GO(Thermo Fisher Scientific);微波消解仪Mars 5(CEM Corporation);电感耦合等离子体发射光谱仪Optima 2000 DV(Perkin Elmer)。
1.2 种子和幼苗处理
挑选大小均匀、籽粒饱满的种子,用70%乙醇浸种2 min,再用3%H2O2消毒2次,然后使用蒸馏水冲洗干净。将种子整齐排列在铺有滤纸的培养皿中,于25 ℃恒温培养箱培养。待种子露白后转入培养液中,在光照培养箱中继续培养,8:00—20:00温度控制在25 ℃,20:00至次日8:00温度控制在18 ℃。当幼苗长到2叶1心时进行盐胁迫处理。
1.3 水培试验
1.3.1 试验设计
利用Nipponbare水稻品种进行水培试验,当幼苗长到2叶1心时进行盐胁迫处理,试验设3个处理,分别为营养液组(CK),0.5% NaCl营养液组和3 mmol/L Na2SiO3+ 0.5% NaCl营养液组,每个处理设3个重复。培养皿中的盐溶液每3 d更换1次。幼苗在盐胁迫处理3 和7 d后,每组取10株测量生长指标;盐胁迫处理7 d后,观察其盐害症状并进行生理指标测定。
1.3.2 盐害症状观察
盐胁迫处理7 d,记录不同处理下,仅第1片叶发黄(2级)、第1、2片叶发黄卷曲(4级)、第3片叶叶尖卷曲发黄(6级)、第3片叶全部发黄卷曲(8级)和叶片全部枯死(10级)的水稻幼苗数量。生长发育正常,不表现任何盐害症状记为0级。水稻幼苗盐害指数计算如式(1) 。
盐害指数(%)=∑[(各级受害植株数×相应级数值)/(总株数×最高盐害级数值)]×100
1.3.3 生长指标测定
在盐处理3和7 d后分别测量幼苗的苗长,根长,根、茎叶鲜重和干重及第3叶叶长等生长指标。苗长:茎基部至植株顶端生长点。根长:茎基部至最长根尖。第3叶片叶长:第3片完全展开叶从叶鞘基部至叶尖的长度。将幼苗分为根系和茎叶两部分,用吸水纸吸干表面水分后,采用精度为0.001 g的电子天平称量根鲜重和茎叶鲜重,随后将样品置于烘箱中,经105 ℃杀青30 min、75 ℃烘至恒重,冷却至室温后称量根干重和茎叶干重。
1.3.4 生理生化指标测定
盐处理7 d后,采集幼苗植株,将幼苗分为根和茎叶两部分,测定植株不同部位的CAT、POD、SOD、APX活性和MDA含量。(1)CAT、POD、SOD、APX活性检测。准确称取植物组织,按质量体积比1∶9的比例加入生理盐水,冰浴条件下,制备10%的组织匀浆,2 500×g,离心10 min,取上清液后用生理盐水稀释成最佳浓度,用酶标仪测定吸光度值;具体操作方法参照试剂盒说明书。(2)MDA含量测定。准确称取植物组织,按质量体积比1∶9的比例加入MDA含量测定试剂盒中的试剂5提取液,将样品剪碎后用内切式匀浆机冰水浴匀浆,离心后取上清液,用酶标仪测定吸光度值;具体方法参照试剂盒说明书。
1.4 田间盆栽试验
1.4.1 试验设计
利用连粳7号和连粳15号水稻进行盆栽试验。设置4种育秧基质处理,分别为育秧基质(对照)、育秧基质+Na2SiO3、育秧基质+1/2硅藻土、育秧基质+1/4硅藻土。水稻幼苗长至2叶1心时,进行盐胁迫(0.5% NaCl)处理。
1.4.2 幼苗生长情况及盐离子浓度测定
盐胁迫7 d后,观察各处理下幼苗生长情况,测定分析水稻幼苗体内的盐离子浓度及分布。首先将水稻茎叶和根分别磨碎,称取0.1 g左右的样品放入干燥洁净的微波硝化罐中,加入2.5 mL浓硝酸,放入微波消煮炉中,90 ℃消解50 min,硝化好的样品用超纯水定容至25 mL,过滤后用电感耦合等离子体发射光谱仪进行盐离子浓度测定。
1.5 数据处理
采用 GraphPad Prism 软件进行数据处理。
2 结果与分析
2.1 水稻幼苗盐害情况
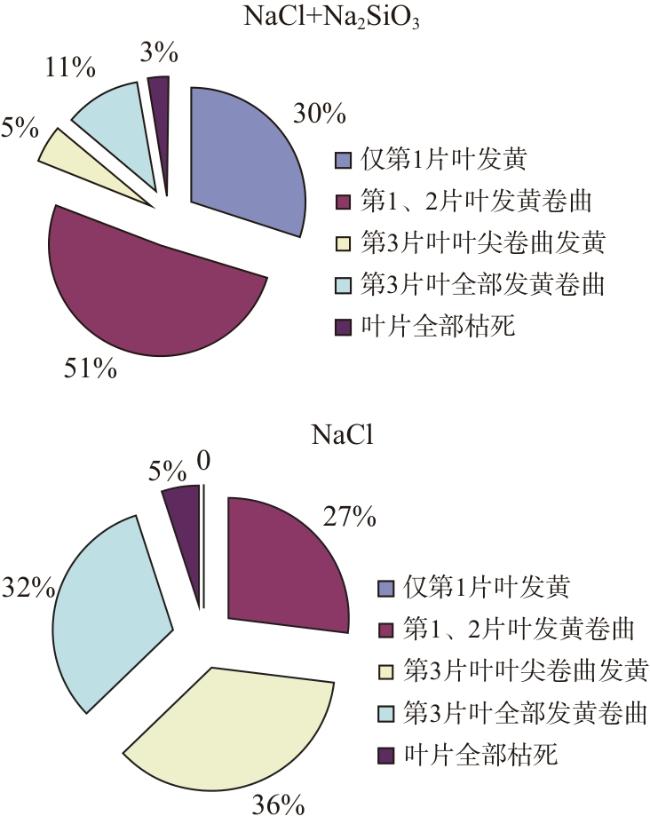
由图1可知,NaCl处理7 d后,水稻幼苗均出现叶片发黄或卷曲现象,且发黄卷曲叶片均在2片及以上,第3片叶出现盐害反应的比例为68%,水稻幼苗死亡率为5%。在NaCl+Na2SiO3处理下,水稻死亡率为3%,30%的水稻幼苗仅第1、2片叶发黄卷曲,第3片叶发黄卷曲的比例为16%,与单独盐处理相比降低了52个百分点。NaCl处理下水稻幼苗盐害指数为63.0%,NaCl+Na2SiO3处理下水稻幼苗盐害指数为41.2%。说明在盐胁迫环境下,添加外源硅可有效缓解水稻幼苗的盐害症状,降低其盐害指数。
2.2 水稻幼苗生长指标
2.2.1 盐胁迫处理3 d
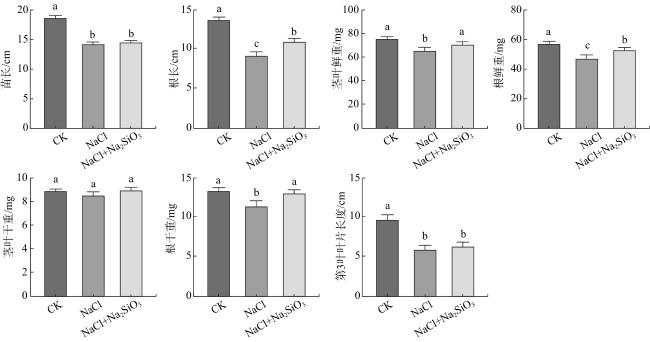
盐胁迫处理3 d后,各处理的茎叶干重无明显变化;NaCl处理的幼苗苗长、根长、茎叶鲜重、根鲜重、根干重和第3叶叶长均低于CK,差异具有统计学意义(P<0.05);而NaCl+Na2SiO3处理的根长、茎叶鲜重、根鲜重和根干重均高于NaCl处理,差异具有统计学意义(P<0.05)(图2)。
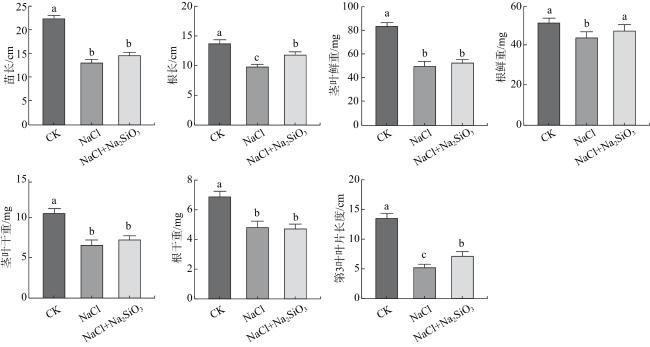
2.2.2 盐胁迫处理7 d
2.3 生理生化指标
2.3.1 酶活性指标
由图4可知,水稻幼苗盐处理7 d后,叶片中NaCl处理的CAT活性较CK降低了82.4%,差异具有统计学意义(P<0.05);NaCl+Na2SiO3处理的CAT活性较NaCl处理提高了65.4%,差异具有统计学意义(P<0.05)。NaCl处理的SOD活性与CK差异无统计学意义(P>0.05);NaCl+Na2SiO3处理的SOD活性明显低于NaCl处理(P<0.05)。叶片中的POD和APX活性在各处理间差异无统计学意义(P>0.05)。
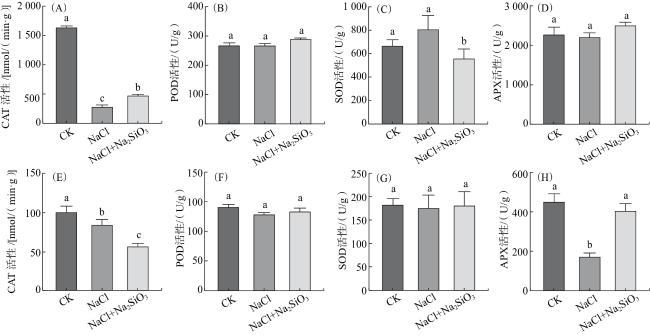
由图4可知,在根部,与CK相比,NaCl处理的 CAT、APX活性均明显降低(P<0.05);NaCl+Na2SiO3处理的APX活性明显高于NaCl处理(P<0.05),CAT活性明显低于NaCl处理(P<0.05)。根部的SOD和POD活性在各处理间差异无统计学意义(P>0.05)。
2.3.2 MDA含量
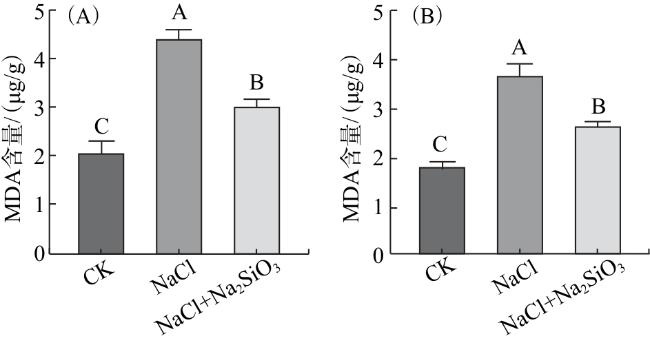
2.4 水稻的生长情况


2.5 水稻幼苗植株内离子分布
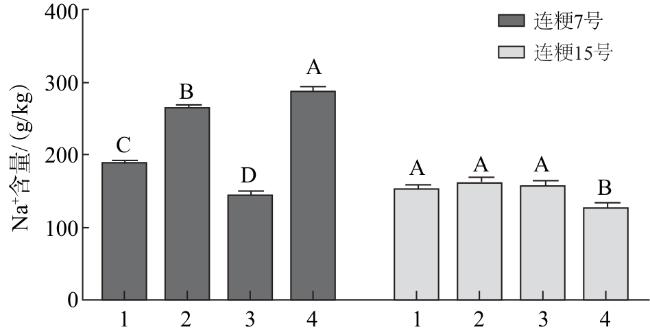
由图8可知,连粳7号整株Na+含量(根部+地上部)高于连粳15号。在连粳7号的外源硅处理中,除1/2硅藻土处理外,其余植株Na+含量均高于NaCl处理;在连粳15号中,植株Na+含量除1/4硅藻土处理略有降低外,其余处理含量变化不明显。
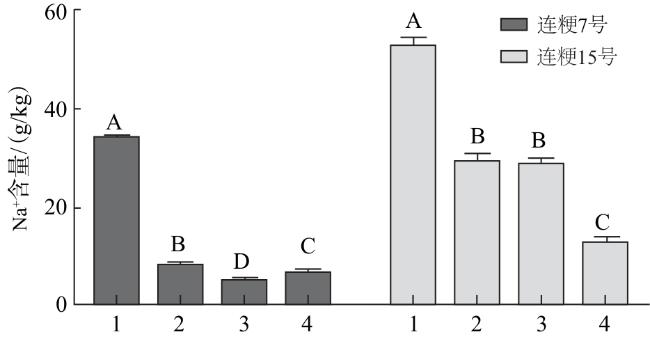
由图9可知,连粳7号根部Na+含量低于连粳15号。与NaCl处理相比,添加外源硅后(Na2SiO3、硅藻土),水稻幼苗根部的Na+含量均明显降低(P<0.01);其中连粳7号添加1/2硅藻土后Na+含量降低的最多;连粳15号添加1/4硅藻土后Na+含量降低的最多。
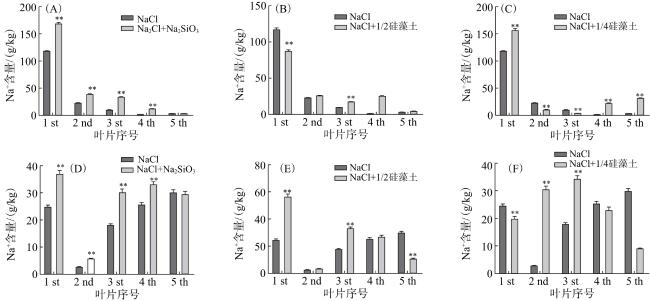
由图10可知,2个品种在Na2SiO3处理中,除新叶(第5片叶)外,其他叶片的Na+含量均较NaCl处理明显增加。连粳7号Na+主要分布在第1叶片(老叶中);连粳15号大部分叶片分布较均匀。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 结论与讨论
水稻幼苗期对盐浓度较为敏感,探索幼苗在盐胁迫下的安全性是当前水稻抗逆育种工作中亟待解决的问题之一。盐胁迫主要通过破坏植物细胞质膜的完整性、诱导活性氧产生,从而引发膜脂过氧化作用,同时造成植株吸水受阻、体内离子失衡与水分亏缺,进而导致幼苗代谢紊乱,影响其正常生长发育[9]。本研究发现,水稻幼苗在0.5% NaCl处理3和7 d后,其苗长、根长、茎叶鲜重、根鲜重、根干重和第3叶叶长均低于对照,而NaCl+Na2SiO3处理明显提高了幼苗的根长、根鲜重。说明盐胁迫对水稻幼苗的生长具有一定的抑制作用,而添加外源硅(Na2SiO3)在盐胁迫下对幼苗根部生长有明显促进作用,可有效缓解盐离子对根系细胞的渗透胁迫与离子毒害,进而保证根部正常的代谢活动,使营养物质有效地输送到地上部,为幼苗生长发育提供物质保障。
在盐胁迫下,细胞质膜是细胞最先受盐离子胁迫影响的结构。本研究结果显示,在盐胁迫下,水稻幼苗茎叶和根部的MDA含量明显上升,加入Na2SiO3后MDA含量较盐处理有所下降。说明外源硅对植株细胞膜的完整性具有一定的保护作用。此外,植株体内活性氧防御系统对清除活性氧,维持系统平衡具有重要作用。Lekklar等[10]研究表明,盐胁迫下,外源硅的添加增强了CAT活性,且APX活性在叶片和根部均有所提高。Das等[11]研究证明,硅处理显著提高了感盐水稻品种的APX活性。本研究中,与CK相比,单独盐胁迫的水稻叶片中CAT活性明显降低;施加Na2SiO3后,CAT活性较单独盐胁迫明显提高。单独盐处理下,水稻根部CAT、APX活性明显低于CK;加入Na2SiO3后其根部APX活性较单独盐处理明显提高。由此可见,叶片中的CAT活性和根部的APX活性对提升水稻植株应对盐胁迫的抗氧化防御能力具有重要作用。
高盐胁迫下的渗透调节对细胞内许多功能蛋白或酶类的活性至关重要[12]。关于外施硅提升水稻耐盐性的作用机制,目前存在2种观点:一是在盐胁迫下,施硅可降低水稻植株的蒸腾作用强度,从而减少Na+和Cl-的吸收和运输[13];二是硅元素可沉积于水稻根部外皮层细胞和内皮层细胞中,通过阻断盐分在质外体途径的运输路径,进而抑制盐离子在维管束木质部的富集与累积[14]。本研究表明,外源硅减少了盐胁迫下植株根部Na+含量,增加了植株地上部Na+含量。此结果与Munns等[15]的研究结论(将Na+聚集在根部)不一致,可能是因为硅对水稻K+、Na+的吸收和分配与水稻品种基因型有关[8]。闫国超等[8]研究发现,硅在盐胁迫下提高了根部的OsSOS1(Na+外排基因)基因的表达量,这与本研究结果中根部Na+含量减少一致。
综上,本研究通过水培试验和田间盆栽试验,探究了外源硅增强水稻幼苗耐盐性的机理,结果表明,外源硅可通过调控水稻幼苗体内抗氧化酶(CAT、APX)活性,增强植株抗氧化防御能力以减轻盐胁迫造成的氧化损伤;通过调控盐离子在植株体内的分布,以提高水稻幼苗对Na+的耐受性。此外,不同来源的外源硅均可提高水稻幼苗的耐盐能力,缓解盐胁迫,其在盐碱地水稻实际生产中具有较大应用潜力。