

马占相思(Acacia mangium)是华南地区常见的人工林树种,其具有生长迅速、树冠茂盛、固氮能力强和耐瘠薄等特点,在自然分布区内长期处于野生状态,能快速占据可供更新的荒山或采伐迹地,在瘠薄土地上也能生长成林[1-2]。该树种在强化生态防护以及保护林木种质资源等方面发挥着重要作用[2-3]。
水分是干旱、半干旱区植物生长繁殖的主要限制因素之一,其变化对植物生理、生化过程和生态系统结构功能的影响较大[4]。干旱胁迫下,植物体内会发生一系列复杂的生理生化变化,通过调控抗氧化酶系统活性、维持细胞膜系统稳定性、积累渗透调节物质等方式,构建应对外界干旱逆境的适应与抵御机制[5]。叶片形态变化是植物遭受干旱胁迫伤害最直观的外在表现。干旱胁迫对植物的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等光合生理指标均会产生明显影响。研究表明,干旱条件下,植物通过调控渗透调节物质含量和抗氧化相关酶的活性,降低胞内水势,维持细胞内水分平衡,减少活性氧(ROS)积累,从而降低脂质过氧化产物丙二醛(MDA)含量[4,6]。
何琴飞等[1]研究表明,低温胁迫下,相对电导率、游离脯氨酸(Pro)、可溶性蛋白(SP)和可溶性糖(SS)含量可作为相思树种抗寒性评价的参考指标;结合生理指标的变化情况,相思树发生寒害的温度主要在0~6 ℃。杨舒贻等[7]采用水培法测定大叶相思幼苗在不同强度酸胁迫下叶片MDA含量,Pro含量以及过氧化物酶(POD)、谷胱甘肽-S-转移酶(GST)、谷胱甘肽过氧化物酶(GPX)和谷胱甘肽还原酶(GR)等抗氧化活性,结果表明,基于不同梯度酸胁迫下各生理指标的变化规律,推测大叶相思幼苗耐强酸胁迫的生理调节阈值可能为pH 3.0。目前关于马占相思抗旱性评价指标的筛选与选择的研究有待进一步系统完善。
本研究采用盆栽干旱胁迫法,测定干旱胁迫7、14、21、28 d下马占相思的生长指标、抗氧化酶活性、渗透调节物质含量及光合参数等生理生化指标变化,系统探讨马占相思的抗旱生理机制,为其人工栽培、抗逆管理及推广种植提供依据。
1 材料与方法
1.1 试验材料
试验材料为广州花木苗圃基地中两年生马占相思幼苗20株,采用3加仑中号花盆移植,每盆2株,共移植10盆;每盆盛土8 L,土壤使用泥炭土(土壤有机质含量为70%,pH 5.5,电导率为0.5)。正常生长1个月后,选择生长状态良好,长势基本一致的6盆植株开展干旱胁迫试验。
1.2 试验设计
试验地点设在仲恺农业工程学院种植大棚,在胁迫开始前1 d对植株进行充分浇水。试验采用盆栽干旱法,3盆作为对照组(CK组),其间正常养护;另外3盆为干旱处理组(EP组),实施持续干旱胁迫处理,胁迫周期为28 d。干旱胁迫期间,每7 d进行1次植株表型观测、光合参数测定及样本采集,测定时间统一控制在9:00—11:00;根据干旱胁迫天数,干旱处理组依次记为EP7d(干旱7 d)、EP14d(干旱14 d)、EP21d(干旱21 d)和EP28d(干旱28 d)。
1.3 测定指标及方法
1.3.1 植物表型观测
在植株后方放置黑色幕布,选择避光遮阴环境对植株进行拍照,详细记录植株整体生长状态、叶片颜色变化及形态特征等情况;同时,使用直尺测量植株高度。
1.3.2 抗氧化酶活性和MDA含量
超氧化物歧化酶(SOD)活性采用WST-1法测定;过氧化氢酶(CAT)活性采用钼酸铵法测定;POD活性采用愈创木酚法测定;MDA含量采用硫代巴比妥酸法测定。
1.3.3 渗透调节物质
Pro含量采用酸性茚三酮方法测定;SS含量采用蒽酮比色法测定;SP含量采用考马斯亮蓝法测定[8]。
1.3.4 光合指标测定
利用Li-6800便携式光合仪(LI-COR,USA)测定植物的Pn、Tr、Ci、Gs,对每株植物的第3~7片叶片挂牌标记进行测定。
1.3.5 叶绿素含量
参考胡秉芬等[9]的试验方法测定植物叶绿素(Chl)、叶绿素a(Chla)和叶绿素b(Chlb)含量,使用紫外分光光度计(北京普析通用仪器有限责任公司)测定各样本分别在波长665、649 nm处的吸光度OD值,计算如式(1 )~(3 )。
Chl=(20.04 OD649+6.10 OD665)×[V/(1 000×W)]
Chla=(13.7 OD665-5.76 OD649)×[V/(1 000×W)]
Chlb=(25.80 OD649-7.6 OD665)×[V/(1 000×W)]
式中,V为提取液体积,mL;W为叶片鲜重,g。
1.4 数据处理
采用SPSS 22.0软件进行相关性分析和主成分分析,采用Excel和Origin软件进行试验数据整理和图表绘制。
2 结果与分析
2.1 对马占相思生长形态的影响
由图1可知,干旱胁迫前,马占相思生长状态良好,植株挺拔,色泽鲜艳,平均株高62.32 cm。胁迫7 d时,植株底部叶片出现枯黄,叶片萎蔫枯黄比例小于10%,株高66.53 cm,较CK组增加6.76%;胁迫14 d时,新叶变薄、中部叶片萎蔫下垂,底部叶片凋落,枯萎程度10%~20%,株高69.43 cm,较EP7d组增加4.36%;胁迫21 d时,所有叶片薄化下垂,新叶往下焦化干枯,50%叶片凋亡,株高70.83 cm,较EP14d组增加2.02%;胁迫28 d时,植株干化、叶片枯萎下垂,底部叶片80%凋亡,株高69.87 cm,较EP21d组减少1.36%,植株干枯濒临死亡。随着干旱胁迫时间的延长,植株生长受到显著抑制,株高增幅逐渐降低;至胁迫后期,植株基本停止生长,且出现叶片萎蔫、皱缩等受害症状。

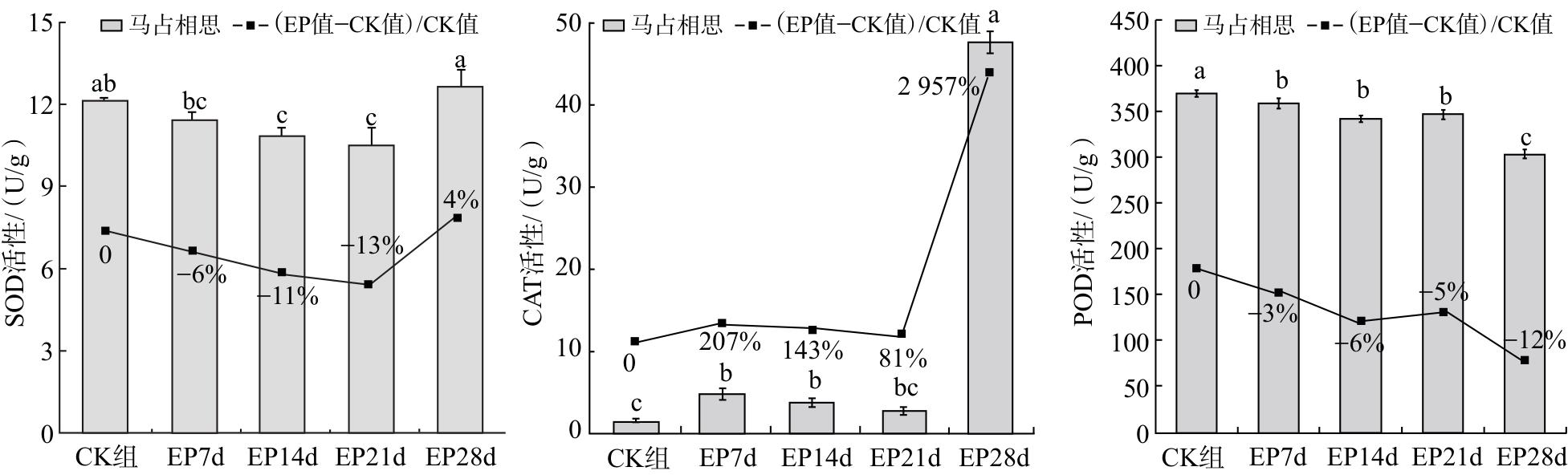
2.2 对马占相思幼苗抗氧化酶的影响
由图2可知,随着干旱胁迫时间的延长,马占相思幼苗的SOD活性呈先下降后上升的趋势,CAT活性呈先上升后下降再上升的趋势,POD活性呈先下降后上升再下降的趋势。其中,干旱胁迫21 d时,马占相思幼苗SOD活性降至最低,与CK组差异具有统计学意义(P<0.05);胁迫末期SOD活性较上一阶段显著增强(P<0.05)。干旱胁迫7 d时,幼苗CAT活性较CK组显著升高(P<0.05);干旱胁迫28 d时,CAT活性明显升高,较CK组上升2 957%。POD活性在干旱胁迫前3个阶段均无明显变化,至胁迫末期较CK组显著降低12%(P<0.05)。
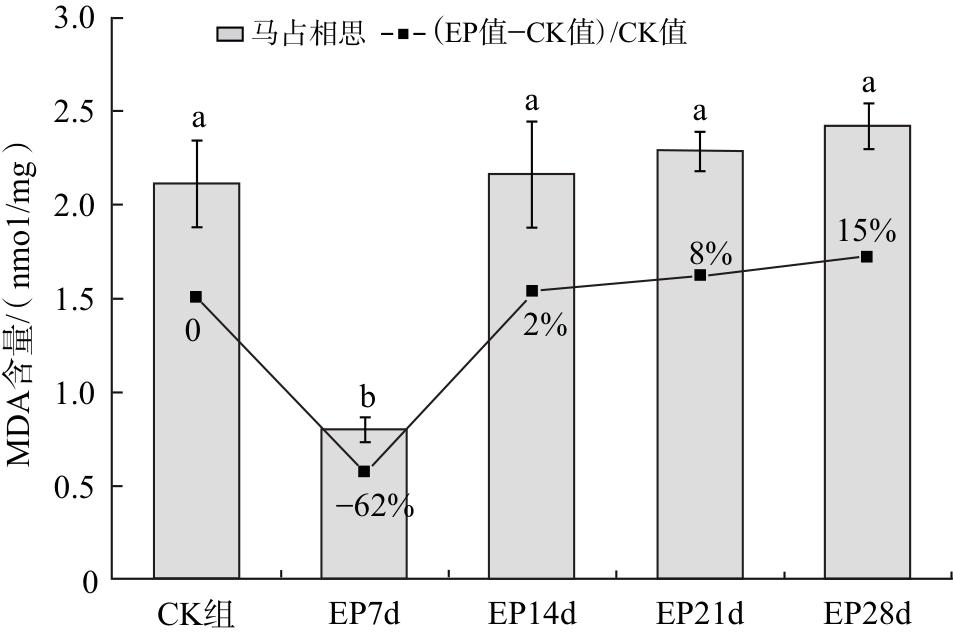
2.3 对马占相思幼苗MDA含量的影响
随着干旱胁迫时间的延长,MDA含量呈先下降后上升的趋势。干旱胁迫7 d时,马占相思幼苗MDA含量显著下降至最低值,较CK组减少62%,胁迫14~28 d期间,幼苗MDA含量较CK组差异无统计学意义(P<0.05),至胁迫末期时,叶片中MDA含量较CK组增加15%(图3)。
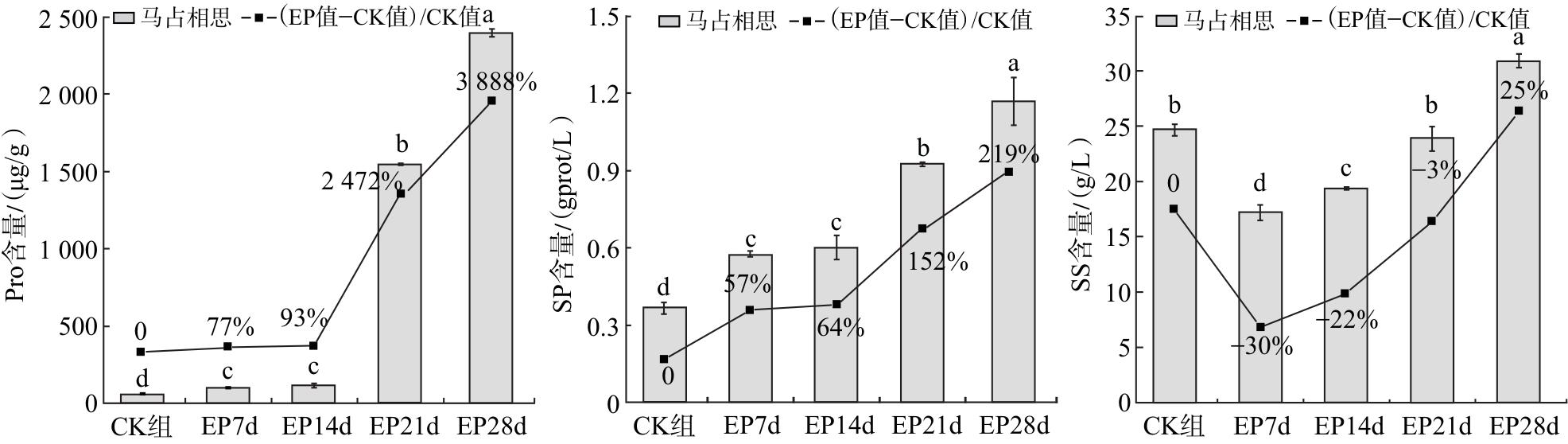
2.4 对马占相思幼苗渗透调节物质的影响
由图4可知,随着干旱胁迫时间的延长,马占相思叶片中Pro和SP含量呈增加的趋势,SS含量呈先下降后上升的趋势。干旱胁迫21和28 d时,Pro含量较CK组上升2 472%和3 888%,差异具有统计学意义(P<0.05);干旱胁迫28 d时,SP含量较CK组增加219%。马占相思叶片SS含量在干旱胁迫7 d时显著下降,降幅达30%(P<0.05);随后随胁迫时间延长逐渐回升,至21 d时与CK组差异无统计学意义(P>0.05);至28 d时显著高于CK组,增幅为25%(P<0.05)。
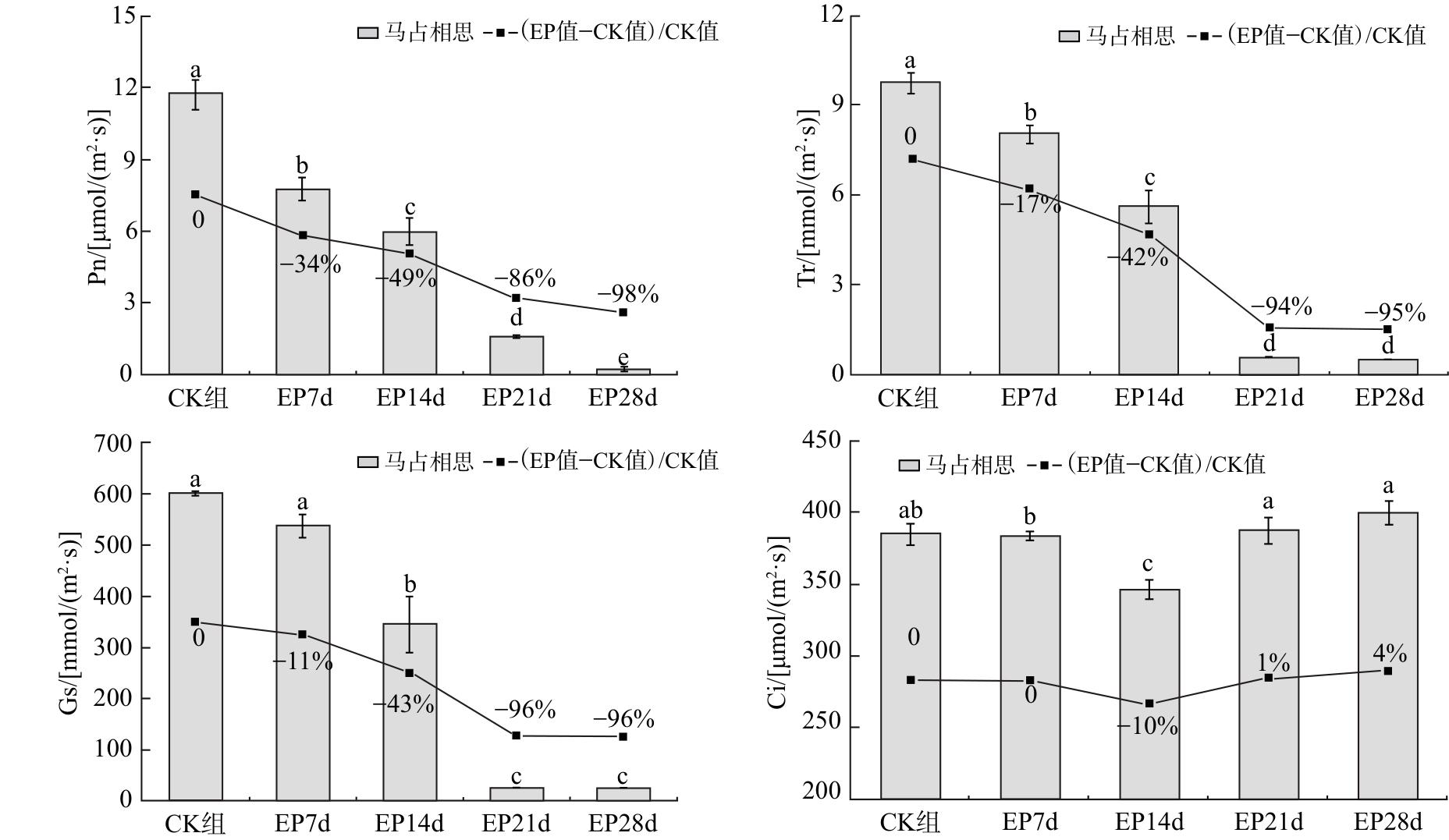
2.5 对马占相思叶片光合参数的影响
由图5可知,随着干旱胁迫时间的延长,马占相思叶片的Pn、Tr和Gs均呈下降趋势,干旱胁迫28 d时,三者较CK组分别下降98%、95%和96%;Ci表现为先下降后上升的趋势,Ci含量在干旱胁迫14 d时最低,较CK组明显下降10%(P<0.05),干旱胁迫21和28 d含量较CK组分别上升1%和4%。
2.6 对马占相思叶片叶绿素含量的影响
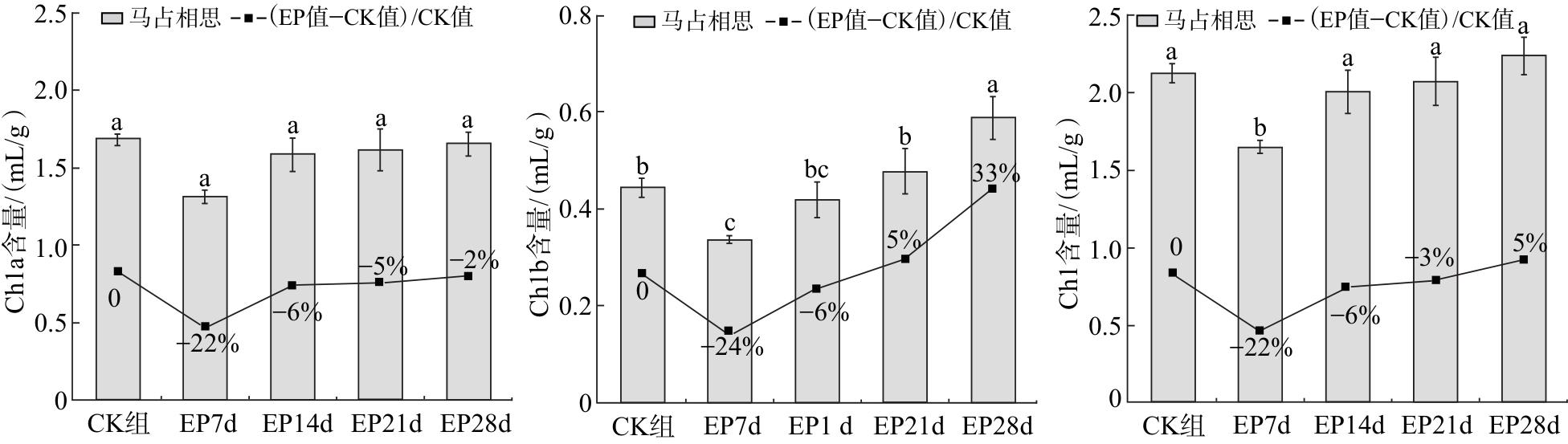
由图6可知,随着干旱胁迫时间的延长,马占相思叶片中的Chl、Chla和Chlb含量均呈先下降后上升的趋势。干旱胁迫7 d时,Chl、Chla和Chlb含量均下降至最低值,此后,3 种叶绿素含量随干旱胁迫时间延长呈逐步上升趋势。干旱胁迫28 d时,Chl和Chlb含量较CK组分别增加5%和33%,Chla含量较CK组降低2%。在干旱胁迫期间,各处理组Chla含量差异无统计学意义(P>0.05)。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.7 干旱胁迫下马占相思抗旱性综合评价
2.7.1 相关性分析
对马占相思在干旱胁迫期间15项生长和生理生化指标进行相关性分析,分析结果见表1。其中,POD活性与SP含量呈显著负相关(P<0.05),与Pn呈显著正相关(P<0.05);MDA含量与Chl、Chla含量均呈显著正相关(P<0.05);Pro含量与SP含量呈极显著正相关(P<0.01),与Pn、Tr、Gs呈显著负相关(P<0.05);SP含量与Pn呈极显著负相关(P<0.01),与Tr、Gs呈显著负相关(P<0.05);SS含量与Chlb含量呈极显著正相关(P<0.01);Chl含量与Chla含量呈极显著正相关(P<0.01),与Chlb含量呈显著正相关(P<0.05);Pn与Tr、Gs呈极显著正相关(P<0.01),与株高呈显著负相关(P<0.05);Tr与Gs呈极显著正相关(P<0.01),与株高呈现显著负相关(P<0.05);Gs与株高呈现显著负相关(P<0.05)。
表1 干旱胁迫下马占相思各指标的相关系数矩阵 |
| 指标 | CAT | POD | SOD | MDA | Pro | SP | SS | Chl | Chla | Chlb | Pn | Tr | Ci | Gs | 株高 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAT | 1 | ||||||||||||||
| POD | -0.843 | 1 | |||||||||||||
| SOD | 0.693 | -0.215 | 1 | ||||||||||||
| MDA | 0.357 | -0.442 | 0.120 | 1 | |||||||||||
| Pro | 0.802 | -0.804 | 0.299 | 0.545 | 1 | ||||||||||
| SP | 0.785 | -0.895* | 0.155 | 0.435 | 0.963** | 1 | |||||||||
| SS | 0.773 | -0.552 | 0.648 | 0.732 | 0.813 | 0.651 | 1 | ||||||||
| Chl | 0.505 | -0.432 | 0.389 | 0.957* | 0.602 | 0.445 | 0.876 | 1 | |||||||
| Chla | 0.251 | -0.206 | 0.247 | 0.954* | 0.387 | 0.215 | 0.729 | 0.960** | |||||||
| Chlb | 0.798 | -0.719 | 0.488 | 0.816 | 0.877 | 0.770 | 0.962** | 0.895* | 0.739 | 1 | |||||
| Pn | -0.639 | 0.879* | 0.082 | -0.430 | -0.890* | -0.970** | -0.497 | -0.370 | -0.172 | -0.667 | 1 | ||||
| Tr | -0.582 | 0.816 | 0.106 | -0.556 | -0.908* | -0.949* | -0.569 | -0.489 | -0.320 | -0.728 | 0.981** | 1 | |||
| Ci | 0.521 | -0.156 | 0.577 | 0.025 | 0.625 | 0.476 | 0.637 | 0.241 | 0.078 | 0.480 | -0.292 | -0.317 | 1 | ||
| Gs | -0.575 | 0.808 | 0.102 | -0.605 | -0.907* | -0.938* | -0.594 | -0.536 | -0.376 | -0.753 | 0.970** | 0.998** | -0.303 | 1 | |
| 株高 | 0.363 | -0.776 | -0.407 | 0.453 | 0.677 | 0.804 | 0.248 | 0.289 | 0.170 | 0.482 | -0.923* | -0.916* | -0.064 | -0.912* | 1 |
|
2.7.2 主成分分析
在干旱胁迫过程中,对15项生长、生理生化指标共提取出3个主成分因子,其特征值分别为9.373、2.893和2.009,方差贡献率分别为62.488%、19.286%和13.394%,累计贡献值达到95.168%。因此,可将15项测定指标转化成3个主成分进行分析。PC1中载荷系数较大的3项分别为Pro、Chlb和SP,且均为正载荷,反映出干旱胁迫下马占相思叶片渗透调节物质和Chlb含量变化等信息;PC2中载荷系数较大的指标分别为SOD、株高和Chla,表明该主成分中主要反映马占相思氧离子消耗和生长变化等信息;PC3中载荷系数较大的指标分别为Ci、MDA和Chla,主要反映出干旱胁迫下植株的膜系统和气孔调节的作用(表2)。综合来看,干旱胁迫下马占相思各主成分下的指标载荷系数最大的关键因子分别为Pro、SOD和Ci,载荷系数较大的有SP、Chlb、Tr、Gs、株高、MDA和Chla等因子。因此,马占相思的抗旱能力大小主要由渗透调节物质因子决定,同时受参与光合作用及叶绿素代谢相关因子的协同调控。
表2 干旱胁迫下马占相思各指标的特征向量和贡献率 |
| 指标 | 主成分因子 | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| CAT | 0.801 | 0.183 | 0.453 |
| POD | -0.850 | 0.279 | -0.148 |
| SOD | 0.287 | 0.763 | 0.522 |
| MDA | 0.721 | 0.306 | -0.622 |
| Pro | 0.965 | -0.063 | 0.206 |
| SP | 0.927 | -0.296 | 0.230 |
| SS | 0.850 | 0.516 | 0.060 |
| Chl | 0.748 | 0.514 | -0.419 |
| Chla | 0.559 | 0.553 | -0.615 |
| Chlb | 0.947 | 0.315 | -0.060 |
| Pn | -0.871 | 0.484 | -0.080 |
| Tr | -0.904 | 0.403 | 0.047 |
| Ci | 0.450 | 0.332 | 0.631 |
| Gs | -0.915 | 0.367 | 0.096 |
| 株高 | 0.713 | -0.662 | -0.226 |
| 特征值 | 9.373 | 2.893 | 2.009 |
| 方差贡献率/% | 62.488 | 19.286 | 13.394 |
| 累积贡献率/% | 62.488 | 81.774 | 95.168 |
3 结论与讨论
SOD、CAT和POD三者协同发挥作用,是植物抵御干旱胁迫的重要途径。本研究显示,在干旱胁迫过程中,EP7d处理组马占相思幼苗CAT活性显著升高,表明试验前期马占相思叶片CAT对干旱胁迫响应敏感,可迅速启动应激反应,有效清除体内产生的ROS,抑制膜脂过氧化进程;与此同时,马占相思叶片中脂质过氧化产物MDA含量也随之显著下降,进一步印证了CAT在干旱胁迫前期对植株的保护作用。与胁迫前期相比,胁迫21 d马占相思叶片SOD、CAT活性均降低,至胁迫28 d二者活性显著升高;而POD活性则在整个干旱胁迫过程中持续降低。这一结果表明,马占相思叶片SOD在干旱胁迫前期并未发挥主要的抗氧化保护作用,直至胁迫后期才逐步启动并发挥一定的调控作用;而干旱胁迫对POD活性具有显著的抑制效应,使其无法有效参与ROS清除及抗逆调控过程。随着干旱胁迫的加剧,植物细胞的代谢稳态被打破,细胞发生膜脂质过氧化反应,产生脂质过氧化产物MDA,使细胞膜的流动性以及渗透性发生改变,进而影响细胞正常功能[12]。本研究发现,干旱胁迫7 d时,马占相思幼苗MDA含量显著下降,随后再次增加,但与CK组相比无明显差异,表明干旱胁迫对马占相思幼苗细胞膜影响较小。
Pro、SS和SP是植物体内3种重要的渗透调节物质。植物体内Pro含量在受到逆境胁迫时会大量积累,抗旱性强的植物体内Pro含量增长幅度更显著[13]。本研究中,随着干旱胁迫程度的增强,马占相思叶片Pro含量呈显著上升趋势,干旱末期其含量较CK组增加3 888%,表明Pro含量变化在马占相思的抗旱过程中发挥较大作用。同时,马占相思叶片中SP含量也呈显著上升趋势,可能是在干旱胁迫过程中,马占相思体内与抗旱相关的蛋白发生特异性表达,进而合成大量胁迫诱导蛋白,参与植株抗旱代谢调控[14]。随着干旱胁迫程度的增加,SS含量呈先下降后上升的趋势,EP7d处理组的叶片SS含量下降可能是前期受到失水胁迫,SS在水解酶的作用下参与细胞内糖酵解过程,而后SS含量在胁迫各阶段均显著增加,提高马占相思叶片的渗透调节能力[15]。Pro、SP、SS含量的显著上升降低了叶片细胞渗透势,植株通过调节渗透调节物质,改变细胞膜的选择透过性,以此抵抗胁迫带来的伤害,维持植物体内正常的水分平衡[16]。
干旱胁迫会导致植物部分乃至全部气孔关闭,进而使植株蒸腾速率、CO2吸收量及光合速率显著下降[17]。干旱胁迫下,植物光合作用的抑制效应主要源于气孔限制与非气孔限制。具体而言,当Pn下降且Ci同步降低时,表明光合作用的减弱主要由气孔限制因素导致;而当Pn下降但Ci升高,且Tr、Gs均显著降低时,则说明光合作用主要受制于非气孔限制因素[18]。本研究表明,马占相思叶片内Pn、Tr、Gs均呈下降趋势,表明在干旱胁迫过程中,马占相思的光合作用、蒸腾作用受到抑制,与陈海鹏等[18]、赵瑜琦等[19]的研究结果均一致。本研究中,马占相思叶片Ci随干旱胁迫时间延长呈先降后升的变化趋势。结合光合参数变化可知,在胁迫前期至14 d,光合作用抑制主要由气孔限制因素导致;而后期光合速率的下降,则主要受制于非气孔限制因素。这表明,干旱胁迫后期已造成叶片光合结构受损,其光化学能力受到显著制约。本研究中,干旱胁迫7 d时,马占相思叶片Chl、Chla、Chlb含量较CK组均降低,表明水分胁迫会抑制叶绿素的生物合成,提高叶绿素酶活性并加速分解,导致叶绿素含量下降[20]。此后,随着干旱胁迫程度的增加,叶绿素含量呈上升趋势,表明持续干旱胁迫抑制了马占相思叶片叶绿素的降解。
本研究结合相关性分析和主成分分析,对马占相思的抗旱性指标进行综合评价。结果表明,在应对干旱胁迫时,Pro、SOD和Ci是马占相思响应干旱胁迫较敏感的生理指标,可作为其抗旱性评价的首选参数,载荷系数较大的指标还有SP、Chlb、Tr、Gs、株高、MDA等因子。综上,马占相思的抗旱能力主要由渗透调节物质因子决定,其次由参与光合作用调控及叶绿素代谢相关的因子协同作用,共同维系植株在干旱逆境下的生长与生理平衡。